




| Original Article | ||
Open Vet. J.. 2022; 12(5): 639-648 Open Veterinary Journal, (2022), Vol. 12(5): 639–648 Original Research Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factorDoaa H. Elsayed1*, Seham A. Helmy2,3, Amina A. Dessouki4, Amal M. El-Nahla5, Heba M. A. Abdelrazek5 and Heba Nageh Gad El-Hak61Department of Theriogenology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt 2Department of Nursing, College of Applied Medical Sciences, University of Bisha, Bisha, Saudi Arabia 3Department of Cytology and Histology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt 4Department of Pathology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt 5Department of Physiology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt 6Department of Zoology, Faculty of Sciences, Suez Canal University, Ismailia, Egyp *Corresponding Author: Doaa H. Elsayed. Department of Theriogenology, Faculty of Veterinary Medicine, Suez Canal University, Ismailia, Egypt. Email: maryam_mohamed2001 [at] yahoo.com Submitted: 26/01/2022 Accepted: 10/08/2022 Published: 10/09/2022 © 2022 Open Veterinary Journal
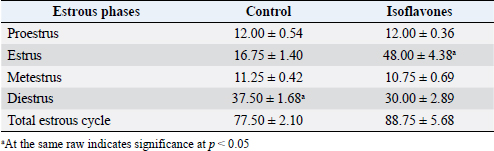
AbstractBackground: Isoflavones are estrogenic compounds that exist in soy, clover, and peanuts. They are selective estrogen receptor modulators. Aim: The study was planned to explain the interactions of isoflavones with estrogen receptors alpha (ERα), beta (ERβ), and vascular endothelial growth factor (VEGF) expressions in ovarian and uterine tissues during different stages of the estrous cycle of regular cyclic female Wistar rats. Methods: Thirty-two regular cyclic females were divided equally into control group: fed casein-based diet and isoflavones group: fed casein-based diet and gavaged 50 mg/kg/day soy isoflavones extract 40%. The regularity of estrus cycles was monitored. Final body weight (FBW), weight gain (BWG), and ovarian and uterine weights were estimated. Histopathology and immunohistochemistry for ERα, Erβ, and VEGF in ovarian and uterine tissues were performed. Results: All females (100%, n=16) in control group showed regularity in estrous cycle compared to 62.5% (n=10) in isoflavones group. Estrus and diestrus phases revealed prolongation and shortening in isoflavones rats than control, respectively. Nonsignificant variation was noted in the duration of the whole cycle of both groups. FBW and BWG significantly decreased however, ovarian and uterine weights increased significantly in all estrous phases of isoflavones group than control. Histopathology demonstrated an increase in number of follicles/ovaries besides, hyperplasia and proliferation of luminal epithelium with hydropic degeneration in the isoflavones group. Also, uterine connective tissue stroma showed edema in the isoflavones group during all estrous phases. Immunostaining percentages of ERα, Erβ, and VEGF protein expression were significantly elevated in the isoflavones group during all estrous phases. Conclusion: Isoflavones induced irregularity of the estrous cycle that was encountered by increased and altered ERα, Erβ, and VEGF expressions in ovarian and uterine tissues. Keywords: ERα, ERβ, Estrous cycle, Isoflavones, VEGF. IntroductionIn females, reproductive capacity is monitored by adjusted reproductive physiology associated with balanced hormonal production, the capability of gametogenesis as well as the morphological and biochemical alterations of the ovary and uterus (Rajan et al., 2017). The estrous cycle is characterized by morphological modifications in ovaries, uterus, and vagina (Goldman et al., 2007). Female rats have a short duration of estrous cycle ranging from 4 to 5 days. Estrous cycle is judged by steroid hormones such as estrogen and progesterone. Estrogen performs its action via binding to estrogen receptors (ERs) and initiating gene expression (da Silva Pacheco et al., 2019). There are two main types of ERs: alpha (ERα) and beta (ERβ). These receptors are extensively spread throughout the reproductive system (Kim and Greenwald, 1987). Selective estrogen receptor modulators (SERMs) are group of nonsteroidal compounds that are capable to bind to ERs prompting alterations in the biological activity of receptors (Xu et al., 2017). In a tissue-specific manner, SERMs proceed with their activities as either agonist or antagonist actions on ERs (Xu et al., 2017). That depends on the involvement of ER signaling which includes the distribution of ERs in different tissues (Wang et al., 2000), ligand binding specificity (Kuiper et al., 1997), and varied interactions with coactivators (Webb et al., 2003). Soy isoflavones are polyphenolic compounds that own a chemical structure analogous to that of estrogen that allows it to bind to nuclear ERs either α and/or β (Franco et al., 2020). Once it binds to ERs, induces receptor-dependent transcription (Morito et al., 2001) by interacting with the estrogen-response element (Patisaul, 2017). Then prompts as an estrogen agonist or antagonist to regulate cell growth, proliferation, and development in the target tissue (Wu et al., 2019). Hence, the biological impacts of isoflavonoids are differed in proportion to the female biological phase (Mc Rodrigues et al., 2018). They also could impair the development of the female genitalia of rodents causing infertility (Nikaido et al., 2004) and implantation losses (Elsayed et al., 2020). Moreover, they were documented to disturb the ovarian function, reduce conception rate as well as upset pregnancy in sheep and cows (Kallela et al., 1984; Adams, 1995). Genistein and daidzein are dominant bioactive members of isoflavones (Šošić-Jurjević et al., 2019). They have antioxidant (Ungar et al., 2003), anti-inflammatory (Wang et al., 2019), and anticancer (Stubert and Gerber, 2009) effects. Isoflavones are thought to be SERMs as they can act with the two subtypes of ERs (Abdelrazek et al., 2019). Vascular endothelial growth factor (VEGF) is an angiogenic factor that promotes vascular permeability (Patel, 2018) and manages the endometrial endothelial cell proliferation (Danastas et al., 2019). It performs its angiogenic role by binding to two tyrosine kinase receptors: VEGFR1 and VEGFR2 which are expressed on the surface of endothelial cells (Terman et al., 1992). It has strong influence on reproductive tract function. It was implicated in implantation (Elsayed et al., 2020), pregnancy (Tousen et al., 2006) as well as cyclic changes (Rimoldi et al., 2007). The influence of soy isoflavones on uterine and ovarian VEGF is debatable, whereas, some studies denoted that they could impede ovarian and uterine VEGF expression (Elsayed et al., 2020). Other studies had shown that isoflavones up-regulate VGEF expression in ovary or uterus (Helmy et al., 2014; Jarić et al., 2018). Moreover, endometrial proliferation, hyperplasia, and neoplasms are common consequences of the uterine ERs up-regulation. Therefore, the elevation of the VEGF expression could be related to endometrial proliferation and neoplasia (Kazi et al., 2005; Newbold et al., 2007; Jarić et al., 2018). The scarcity of literature described the mechanism of isoflavones in cyclic females during the different phases of the estrous cycle. Therefore, this study was designed to elucidate the interactions of isoflavones with ERα, Erβ, and VEGF expressions in ovarian and uterine tissues during different stages of estrous cycle of regular cyclic female Wistar rats with special emphasis on their influence on reproductive cycle regularity. Material and MethodsAnimalsA total of 40 mature female Wistar rats (4.5–5 months; 200–220 g) were used in the study. Animals were purchased from Lab Animal House, Faculty of Sciences, Suez Canal University, Ismailia, Egypt. Rats were housed in plastic cages; five females per cage. They were kept at room temperature (25°C ± 1°C) and natural daylight rhythm. Food and water were offered ad libitum. Rats were adapted for 2 weeks before the beginning of the experiment. Animals were fed with a casein-based diet. Reproductive proceduresAfter 2 weeks of acclimatization, the estrous cycle was monitored by daily cytological examination of vaginal smear according to Singletary et al. (2005). Vaginal smears were obtained daily from each female rat early in the morning. Females with two successive regular cycles were selected for this study, while those of irregular cycles were excluded. Experimental designOnly 32 females exhibited a regular estrous cycle. Regular cyclic rats were divided equally into two groups; G1: control group (n=16) that fed casein-based diet and G2: isoflavones group (n =16) that fed casein-based diet and gavaged 50 mg/kg/day soy isoflavones extract 40% (JMS Vitamins, USA) was dissolved in carboxymethylcellulose with ultrapure water. Each g contained 436 mg isoflavones 132 mg/g genistein and 304 mg/g daidzein that were tested by high-performance liquid chromatography (Elsayed et al., 2020). Estrous regularityVaginal smears were taken from each rat 8 hours apart to determine the average length/hours for each phase of the estrous cycle for three consecutive cycles. Also, the regularity of estrous cycle was monitored for each experimental female. Table 1. Total estrous cycle duration (hours) and different estrous phases (hours).
Body weight, relative ovarian and uterine weightsBody weights were estimated daily. Weight gain (BWG) was also obtained by subtracting the final weight from the initial weight for each experimental animal. After 30 days of treatment, each female rat was sacrificed after exposing it to 5 ml tetrahydrofuran inhalation (98%, Carlo Erba Reagent Co., Italy) in a desiccator. The ovaries and uteri were dissected from each rat. The relative ovarian and uterine weights were recorded by assessment of the weights of ovaries and uteri per gram about body weight. HistopathologyThe dissected ovaries and uteri were immersed in 10% neutral buffer formalin saline. Afterward, they were processed and stained by hematoxylin and eosin (Slaoui and Fiette, 2011). Immunohistochemistry (IHC) and image analysisParaffin-embedded ovaries and uteri were subjected to 5 μm sections used for the IHC of ERα, Erβ, and VEGF as stated by Elsayed et al. (2020). The primary antibodies used from Thermo Scientific Co., (UK) with catalog numbers; # MA1-16629, # RB- 10658-R7, and # MS-750-R7, respectively. The concentrations were 1:50, 1:10, and 1:100, respectively. Image analysis for IHC stained area percentage (ISAP) was done using the Image J program. In all the slides of both groups, nine random fields per slide were randomly selected. Statistical analysisGraphpad prism software (version 7, San Diego, CA) was applied for statistical analysis of the variations between control and soy isoflavones groups in different estrous phases using the Student’s t-test. All the data within the study were expressed as mean ± SEM. Significance was distinguished at a probability value < 0.05. Ethical approvalAll animals were treated and sampled in accordance with the guidelines for care and use of animals which were approved by the research ethics committee in the Faculty of Veterinary Medicine, Suez Canal University (approved no: 2021004 on 27/01/2021).
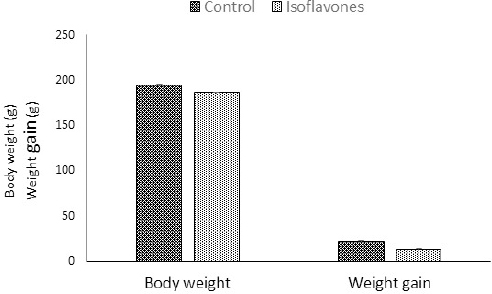
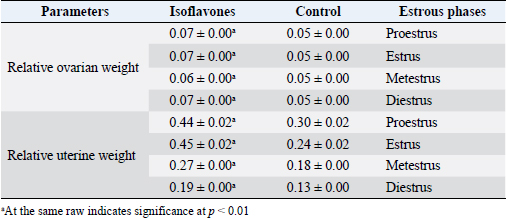
Fig. 1. Body weight (g) and BWG (g) of control and isoflavones cyclic female Wistar rats. Table 2. Relative ovarian (g) and uterine weights (g) of control and isoflavones groups.
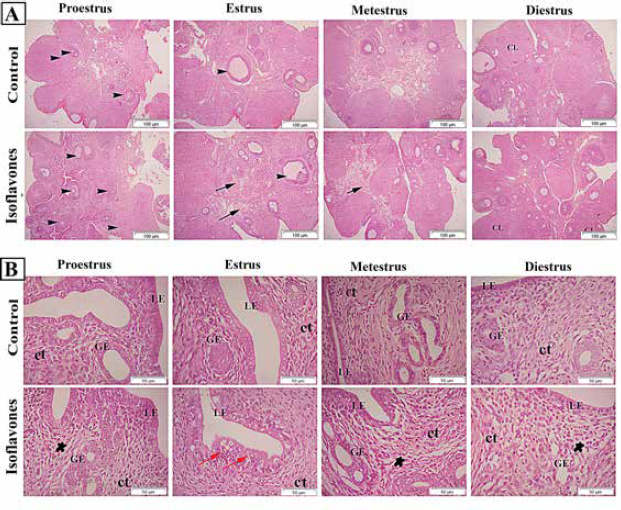
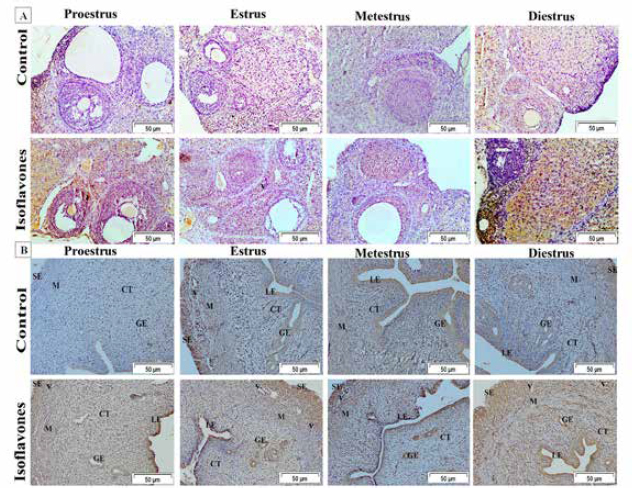
Fig. 2. (A) Photomicrographs in ovary of control and isoflavones groups in cyclic female Wistar rats: ovarian sections showed increased vascularity specially in estrus and metestrus stages (arrows) along with increased number of follicles (arrowhead) in 50 mg/kg/day isoflavones treated group. The figure was stained by H&E (bar 100 µm). (B) Photomicrograph sections in uterus of control and isoflavones groups in cyclic female Wistar rats: uterus sections showed hyperplasia and proliferation of LE with hydropic degeneration (red arrows) of epithelium in estrus phase of 50 mg/kg/day isoflavones treated group. Uterine stromal edema (asterisk) was evident in isoflavones treated groups during all phases of estrous cycle. The figure was stained by H&E (bar 50 µm). ResultsRegularity of estrous cyclesAll females in the control group (100%) showed regularity of estrous cycles. However, 10 (62.5%) out of 16 females in isoflavones group revealed regular estrous cycles. Isoflavones-treated females exhibited significant ( p < 0.0001) prolongation in the duration of estrus phase rather than control. However, the diestrus phase demonstrated significant (p < 0.05) shortening in isoflavones treated rats as compared to control. The proestrus and metestrus durations were nonsignificantly varied between groups. Also, the duration of the whole cycle exhibited a nonsignificant difference between groups (Table 1). Body weight, relative ovarian and uterine weightsSoy isoflavones group revealed significant (p < 0.001) decrease in the final body weight (FBW) and BWG as compared with control group (Fig. 1). However, the relative ovarian and uterine weights in the different estrous phases showed highly significant (p < 0.01) increase in soy isoflavones group as compared to control (Table 2). Ovarian and uterine histopathologyOvary and uterus of control group showed normal histological details related to each phase of estrous. Administration of isoflavones resulted in increment of ovarian and uterine vascularity, especially in estrus and metestrus phases. The number of follicles/ovaries seemed to increase in isoflavones treated group. The estrus phase of isoflavones treated group revealed hyperplasia and proliferation of luminal epithelium (LE) with epithelial hydropic degeneration. Moreover, uterine connective tissue stroma exhibited marked edema in isoflavones treated groups during all phases of estrous cycle (Fig. 2).
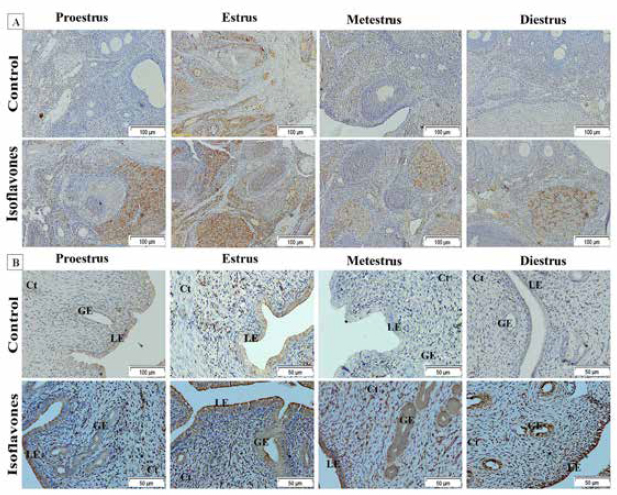
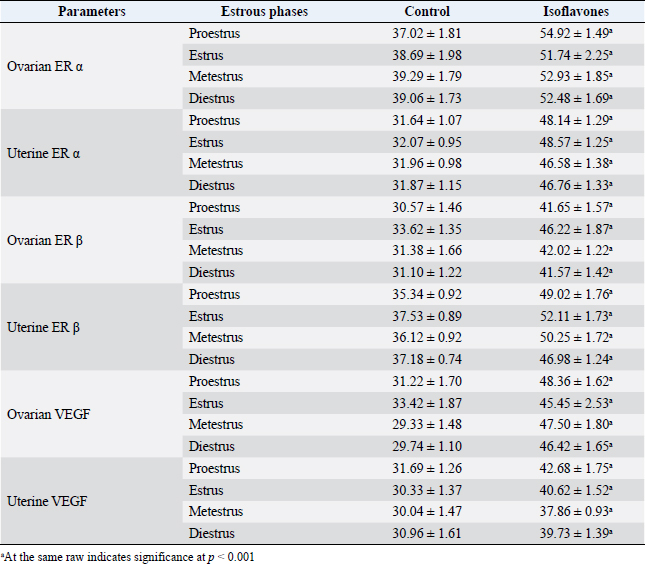
Fig. 3. (A) Photomicrographs of ovarian sections of ERα immunostaining of cyclic female Wistar rats in control and isoflavones groups: positive intracytoplasmic and intranuclear immunoreactivity was pronounced in 50 mg/kg/day isoflavones group in all phases of estrous cycle as compared to control group. The figure was stained by H&E (bar 100 µm). (B) Photomicrographs of uterine sections of ERα immunostaining of cyclic female Wistar rats in control and isoflavones groups: intense intracytoplasmic and intranuclear immunoreactivity was demonstrated in the lining epithelium LE and GE of 50 mg/kg/day isoflavones group rather than control group. The figure was stained by H&E (bar 50 µm). ImmunohistochemistryThe immunostaining of ERα and ERβ were demonstrated as brownish intracytoplasmic as well as intranuclear reactions in ovarian and uterine tissues (Figs. 3 and 4). Ovarian ERα was expressed in the stroma; however, ovarian ERβ was evident in the stroma and corpora lutae of control group. Isoflavones group expressed ERβ in the stroma, corpora lutae and ovarian follicles. Moreover, both receptors were expressed in uterine LE, glandular epithelium (GE), connective tissue stroma (Ct), myometrium (M) and surface epithelium (SE). The ISAP of both ERα and ERβ exhibited significant (p < 0.001) elevations in isoflavones treated group than control during all phases of the estrous cycle (Table 3). The immunostaining of VEGF appeared as brownish intra-cytoplasmic reaction in uterine and ovarian tissues (Fig. 5). The VEGF was expressed in the ovarian stroma as well as corpora lutae. The uterine VEGF signals existed in the cytoplasm of LE, GE, Ct, M, SE, and endothelium of blood vessels (v). The ISAP of VEGF exhibited significant (p < 0.001) elevations in isoflavones treated group than control during all phases of the estrous cycle (Table 3). DiscussionThe effect of isoflavones, as endocrine-disrupting hazard, on the outcome of rodent reproductive performance is controversial (Patisaul, 2017). Isoflavones including; genistein and daidzein are abundant in human and animal diets (Gaffer et al., 2018). Despite the increasing number of studies, there is still a long way to a firm knowledge on the biological potency of dietary isoflavones and their impact on reproduction, especially in mature cyclic females. In this study, the potential estrogenic effects of isoflavones on ERα, Erβ, and VEGF immunohistochemistry were assessed using estrogen-responsive tissues ovary and uterus.
Fig. 4. (A) Photomicrographs of ovarian sections of ERβ immunostaining in control and isoflavones cyclic female Wistar rats: isoflavones group (50 mg/kg/day) showed marked positive intracytoplasmic and intranuclear immunoreactivity especially in all phases of estrous cycle than control group. The figure was stained by H&E (bar 100 µm). (B) Photomicrograph sections of uterus of ERβ immunostaining in control and isoflavones cyclic female Wistar rats: isoflavones group (50 mg/kg/day) revealed positive intracytoplasmic and intranuclear immunostaining reaction in LE, GE and serosa (SE). However, control group showed less immunoreactivity to ERβ in estrous phases. The figure was stained by H&E (bar 100 µm). Soy isoflavones administered group showed a significant reduction in FBW as well as BWG than control. These results were similar to previous results of Elsayed et al., (2020). The reduction in body weight and BWG could be attributed to the appetite repression (Wade, 1975) that is caused by the hypothalamic estrogen stimulating action of isoflavones (Xu et al., 2011). Furthermore, the estrogenic influence of isoflavones could regulate body fats and downgrade their leptin production, therefore, repressing appetite (Szkudelska et al., 2000). It was noticed that soy isoflavones significantly increased both the ovarian and uterine weights during all phases of estrous than control. This increment could be attributed to the increased number of ovarian follicles as well as the observed hyperemia in the histopathological section. Moreover, the increase in the uterine weight represented by edema and water imbibitions in the stroma of isoflavones-treated uteri. This may be attributed to the estrogenic response of isoflavones that also stimulated endometrial proliferation and hyperplasia (Albertazzi and Sharma, 2005). Previous studies have shown similar results and confirmed the estrogenic response of isoflavones in reproductive tract (Rimoldi et al., 2007; Teixeira et al., 2019). On the other hand, Diel et al. (2006) showed that isoflavones did not elicit estrogenic effect or weight variation on reproductive tract. However, Bitto et al. (2010) reported that genistein aglycone might be useful for the management of endometrial hyperplasia in women. Female rats administered soy isoflavones showed alterations in the regularity of the estrous cycle where 10 (62.5%) out of 16 females exhibited irregular cycles. The most common findings were being; prolongation of estrus while shortening the diestrus phase of isoflavones administered group. Others have shown similar estrous cycle alterations in other model systems (Jefferson et al., 2006). Estrogenic potency of isoflavones was reflected in marked changes in vaginal estrous cycle revealing alterations of estrous cyclicity (You et al., 2002) as demonstrated by increased ovarian and uterine ERs ISAP in this study. Isoflavones, especially genistein are more selective to ERβ due to their structural similarity to estrogen (Sirtori et al., 2005). The pattern of estrous cyclicity is usually determined via the circulating level of estrogen and the existence of ERs in both ovarian and uterine tissues. The pattern of their expression could influence estrogen-responsive genes in a manner which is cell specific (Li, 1994). It seems that the disrupted estrous cycle may be attributed to the change in spatial ovarian and uterine ERβ expression. The ERβ is essential to the early-stage follicle growth and the interaction between the two ERs is required for the late stage of follicle growth (Couse et al., 1997). Table 3. Percentages of immunostained area (mean ± SEM) of ovarian and uterine ERα, ERβ and VEGF.
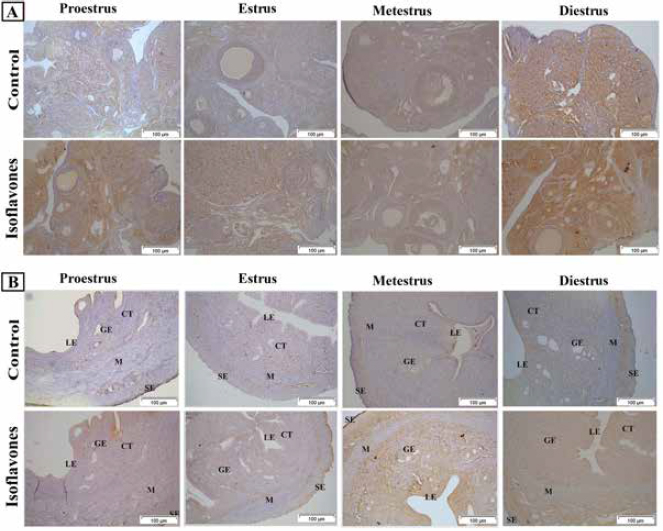
VEGF is expressed in LE, GE, CT stroma, v and M of uterus while in ovary it was expressed in stroma and corpus luteum with higher expression values in isoflavones administered group. This increment in VEGF could be accredited to the estrogenic potential of isoflavones displayed by the promoted uterine cellularity. Some studies displayed the change in expression of VEGF with disparities in ERs concentration (Bausero et al., 1998; Hervé et al., 2006). In any event, VEGF over production probably identifies blood vessels and its size and strongly suggests a role for VEGF in in vivo angiogenesis and microvascular hyperpermeability (Bausero et al., 1998) manifested by edema that was observed in the current study. Moreover, the hyperexpression of VEGF could be involved in pathological situations, abnormal hyperpermeability, and dilated capillaries and increase the risk of uterine cancer (Bausero et al., 1998; Zhang et al., 2016). This suggests that isoflavones could predispose uterine neoplasia. These results are in disagreement with those of Niwa et al. (2000) who demonstrated the inhibitory effect of genistein and daidzein on endometrial carcinogenesis and attributed it to their antiestrogenic action and suppression of its downstream estrogen receptor response elements.
Fig. 5. (A) Photomicrograph sections of ovary of VEGF immunostaining in control and isoflavones cyclic female Wistar rats: estrous phases of 50 mg/kg/day isoflavones group showed more intense positive intracytoplasmic immunoreactivity in all estrous phases. However, control group exhibited less immunoreactivity in VEGF receptors. The figure was stained by H&E (bar 50 µm). (B) Photomicrograph sections of uterus of VEGF immunostaining in control and isoflavone cyclic female Wistar rats: estrous phases of 50 mg/kg/day isoflavones group showed pronounced positive intracytoplasmic VEGF immunoreactive staining. Control group showed less VEGF immunoreactivity than 50 mg/kg/day isoflavones group. The figure was stained by H&E (bar 50 µm). Taking together all the results, it was shown that soy isoflavones as SERM could increase both ERs subtypes. ERα mainly encountered the uterotrophic response as well as VEGF-induced vascular and permeability changes while ERβ encountered cyclic patterns irregularity. ConclusionThe present study showed the adverse effects of isoflavones on the reproductive pattern in cyclic female Wistar rats. Isoflavones induced reproductive cycle irregularities that are encountered by increased and altered ERβ expression pattern. Moreover, proliferative changes were evident that were mediated by ERα expression, especially in uterus. Also, isoflavones up regulated VEGF expression in ERα expression dependent pattern that promoted angiogenesis and hyperpermeability in blood vessels that may predisposes pathological conditions as neoplasia in uterus and ovary. Author’s contributionD.H.E analyzed the data statistically and wrote the manuscript. S.A.H. and A.A.D. prepared the figures. A.M.E. revised the manuscript. H.M.A. and H.N.G performed the study and discussed the results. Conflict of interestAll the authors declare that they have no competing interest. ReferencesAbdelrazek, H.M.A., Mahmoud, M.M.A., Tag, H.M., Greish, S.M., Eltamany, D.A. and Soliman, M.T.A. 2019. Soy isoflavones ameliorate metabolic and immunological alterations of ovariectomy in female Wistar rats: antioxidant and estrogen sparing potential. Oxid. Med. Cell. Longev. 2019, 1–13. Adams, N.R. 1995. Detection of the effects of phytoestrogens on sheep and cattle. J. Anim. Sci. 73, 1509–1515. Albertazzi, P. and Sharma, S. 2005. Urogenital effects of selective estrogen receptor modulators: a systematic review. Climacteric 8, 214–220. Bausero, P., Cavaillé, F., Méduri, G., Freitas, S. and Perrot-Applanat, M. 1998. Paracrine action of vascular endothelial growth factor in the human endometrium: production and target sites, and hormonal regulation. Angiogenesis 2, 167–182. Bitto, A., Granese, R., Triolo, O., Villari, D., Maisano, D., Giordano, D., Altavilla, D., Marini, H., Adamo, E.B., Nicotina, P.A., Anna, R.D., and Squadrito, F. 2010. Genistein aglycone: a new therapeutic approach to reduce endometrial hyperplasia. Phytomedicine 17, 844–850. Couse, J.F., Lindzey, J., Grandien, K., Gustafsson, J.A. and Korach, K.S. 1997. Tissue distribution and quantitative analysis of estrogen receptor-α (ERα) and estrogen receptor-β (ERβ) messenger ribonucleic acid in the wild-type and ERα-knockout mouse. Endocrinology 138, 4613–4621. da Silva Pacheco, S., Rondini, T.A., Bittencourt, J.C. and Fuzeti Elias, C. 2019. Neurons expressing estrogen receptor α differentially innervate the periaqueductal gray matter of female rats. J. Chem. Neuroanat. 97, 33–42. Danastas, K., Whittington, C.M., Dowland, S.N., Combes, V., Murphy, C.R. and Lindsay, L.A. 2019. Ovarian hyperstimulation reduces vascular endothelial growth factor—a during uterine receptivity. Reprod. Sci. 26, 259–268. Diel, P., Hertrampf, T., Seibel, J., Laudenbach-Leschowsky, U., Kolba, S. and Vollmer, G. 2006. Combinatorial effects of the phytoestrogen genistein and of estradiol in uterus and liver of female Wistar rats. J. Steroid. Biochem. Mol. Biol. 102, 60–70. Elsayed, D.H., Abdelrazek, H.M.A., Eltamany, D.A., Ebaid, H.M. and El-Nahla, A.M. 2020. Effect of soy isoflavones on implantation losses in Wistar rat: implication of progesterone receptors, vascular endothelial growth factor and estradiol receptors alpha. Iran. J. Vet. Res. 21, 46. Franco, P.C., Simões, R.S., Carbonel, A.A.F., Sasso, G.R.d.S., Florencio-Silva, R., Baracat, E.C., Girão, M.B.C., Soares Júnior, J.M. and Simões, M.d.J. 2020. The influence of phytoestrogens or estrogens on the proliferation of the rat endocervical mucosa. Rev. Assoc. Med. Bras. 66, 174–179. Gaffer, G.G., Elgawish, R.A., Abdelrazek, H.M.A., Ebaid, H.M. and Tag, H.M. 2018. Dietary soy isoflavones during pregnancy suppressed the immune function in male offspring albino rats. Toxicol. Rep. 5, 296–301. Goldman, J.M., Murr, A.S. and Cooper, R.L. 2007. The rodent estrous cycle: characterization of vaginal cytology and its utility in toxicological studies. Birth. Defects. Res. B. Dev. Reprod. Toxicol. 80, 84–97. Helmy, A., Emarah, H. and Abdelrazek, H. 2014. Estrogenic effect of soy phytoestrogens on the uterus of ovariectomized female rats. Clin. Pharmacol. Biopharm. 2, 7. Hervé, M.A.J., Meduri, G., Petit, F.G., Domet, T.S., Lazennec, G., Mourah, S. and Perrot-Applanat, M. 2006. Regulation of the vascular endothelial growth factor (VEGF) receptor Flk-1/KDR by estradiol through VEGF in uterus. J. Endocrinol. 188, 91–99. Jarić, I., Živanović, J., Miler, M., Ajdžanović, V., Blagojević, D., Ristić, N., Milošević, V. and Nestorović, N. 2018. Genistein and daidzein treatments differently affect uterine homeostasis in the ovary-intact middle-aged rats. Toxicol. Appl. Pharmacol. 339, 73–84. Jefferson, W.N., Padilla-Banks, E. and Newbold, R.R. 2006. Studies of the effects of neonatal exposure to genistein on the developing female reproductive system. J. AOAC. Int. 89, 1189–1196. Kallela, K., Heinonen, K. and Saloniemi, H. 1984. Plant oestrogens; the cause of decreased fertility in cows. a case report. Nord. Vet. Med. 36, 124–129. Kazi, A.A., Jones, J.M. and Koos, R.D. 2005. Chromatin immunoprecipitation analysis of gene expression in the rat uterus in vivo: estrogen-induced recruitment of both estrogen receptor α and hypoxia-inducible factor 1 to the vascular endothelial growth factor promoter. Mol. Endocrinol. 19, 2006–2019. Kim, I. and Greenwald, G.S. 1987. Estrogen receptors in ovary and uterus of immature hamster and rat: effects of estrogens. Endocrinol. Japonica. 34, 45–53. Kuiper, G.G., Carlsson, B., Grandien, K., Enmark, E., Häggblad, J., Nilsson, S. and Gustafsson, J.A. 1997. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 138, 863–870. Li, S. 1994. Relationship between cellular DNA synthesis, PCNA expression and sex steroid hormone receptor status in the developing mouse ovary, uterus and oviduct. Histochemistry 102, 405–413. Mc Rodrigues, G., Db Borges, B., Moreira, L.G.Q., Rossete, É.A.G. and de Castro Franca, S. 2018. Effects of estrogen-like plant compounds on the vaginal epithelium pituitary, adrenal glands, and uterus of rats. Exp. Biol. Med. (Maywood). 243, 1173–1184. Morito, K., Hirose, T., Kinjo, J., Hirakawa, T., Okawa, M., Nohara, T., Ogawa, S., Inoue, S., Muramatsu, M. and Masamune, Y. 2001. Interaction of phytoestrogens with estrogen receptors α and β. Biol. Pharm. Bull. 24, 351–356. Newbold, R.R., Jefferson, W.N., Grissom, S.F., Padilla-Banks, E., Snyder, R.J. and Lobenhofer, E.K. 2007. Developmental exposure to diethylstilbestrol alters uterine gene expression that may be associated with uterine neoplasia later in life. Mol. Carcinog. 46, 783–796. Nikaido, Y., Yoshizawa, K., Danbara, N., Tsujita-Kyutoku, M., Yuri, T., Uehara, N. and Tsubura, A. 2004. Effects of maternal xenoestrogen exposure on development of the reproductive tract and mammary gland in female CD-1 mouse offspring. Reprod. Toxicol. 18, 803–811. Niwa, H., Miyazaki, J. and Smith, A.G. 2000. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat. Genet. 24, 372–376. Patel, S. 2018. Polycystic ovary syndrome (PCOS), an inflammatory, systemic, lifestyle endocrinopathy. J. Steroid. Biochem. Mol. Biol. 182, 27–36. Patisaul, H.B. 2017. Endocrine disruption by dietary phyto-oestrogens: impact on dimorphic sexual systems and behaviours. Proc. Nutr. Soc. 76, 130–144. Rajan, R.K., Kumar, S.S.M. and Balaji, B. 2017. Soy isoflavones exert beneficial effects on letrozole-induced rat polycystic ovary syndrome (PCOS) model through anti-androgenic mechanism. Pharma. Biol. 55, 242–251. Rimoldi, G., Christoffel, J., Seidlova-Wuttke, D., Jarry, H. and Wuttke, W. 2007. Effects of chronic genistein treatment in mammary gland, uterus, and vagina. Environ. Health. Perspect. 115, 62–68. Singletary, S.J., Kirsch, A.J., Watson, J., Karim, B.O., Huso, D.L., Hurn, P.D. and Murphy, S.J. 2005. Lack of correlation of vaginal impedance measurements with hormone levels in the rat. J. Am. Assoc. Lab. Anim. Sci. 44, 37–42. Sirtori, C.R., Arnoldi, A. and Johnson, S.K. 2005. Phytoestrogens: end of a tale? Ann. Med. 37, 423–438. Slaoui, M. and Fiette, L. 2011. Histopathology procedures: from tissue sampling to histopathological evaluation. Methods. Mol. Biol. 691, 69–82. Šošić-Jurjević, B., Lütjohann, D., Renko, K., Filipović, B., Radulović, N., Ajdžanović, V., Trifunović, S., Nestorović, N., Živanović, J., Stojanoski, M.M., Kӧhrle, J. and Milošević, V. 2019. The isoflavones genistein and daidzein increase hepatic concentration of thyroid hormones and affect cholesterol metabolism in middle-aged male rats. J. Steroid. Biochem. Mol. Biol. 190, 1–10. Stubert, J. and Gerber, B. 2009. Isoflavones–mechanism of action and impact on breast cancer risk. Breast. Care. 4, 22–29. Szkudelska, K., Nogowski, L. and Szkudelski, T. 2000. Genistein affects lipogenesis and lipolysis in isolated rat adipocytes. J. Steroid. Biochem. Mol. Biol. 75, 265–271. Teixeira, C.P., Florencio-Silva, R., Sasso, G.R.S., Carbonel, A.A.F., Simões, R.S. and Simões, M.J. 2019. Soy isoflavones protect against oxidative stress and diminish apoptosis in ovary of middle-aged female rats. Gynecol. Endocrinol. 35, 586–590. Terman, B.I., Dougher-Vermazen, M., Carrion, M.E., Dimitrov, D., Armellino, D.C., Gospodarowicz, D. and Böhlen, P. 1992. Identification of the KDR tyrosine kinase as a receptor for vascular endothelial cell growth factor. Biochem. Biophys. Res. Commun. 187, 1579–1586. Tousen, Y., Umeki, M., Nakashima, Y., Ishimi, Y. and Ikegami, S. 2006. Effects of genistein, an isoflavone, on pregnancy outcome and organ weights of pregnant and lactating rats and development of their suckling pups. J. Nutr. Sci. Vitaminol. 52, 174–182. Ungar, Y., Osundahunsi, O.F. and Shimoni, E. 2003. Thermal stability of genistein and daidzein and its effect on their antioxidant activity. J. Agric. Food Chem. 51, 4394–4399. Wade, G.N. 1975. Some effects of ovarian hormones on food intake and body weight in female rats. J. Comp. Physiol. Psychol. 88, 183–193. Wang, A., Wei, J., Lu, C., Chen, H., Zhong, X., Lu, Y., Li, L., Huang, H., Dai, Z. and Han, L. 2019. Genistein suppresses psoriasis-related inflammation through a STAT3-NF-κB-dependent mechanism in keratinocytes. Int. Immunopharmacol. 69, 270–278. Wang, H., Eriksson, H. and Sahlin, L. 2000. Estrogen receptors alpha and beta in the female reproductive tract of the rat during the estrous cycle. Biol. Reprod. 63, 1331–1340. Webb, P., Nguyen, P. and Kushner, P.J. 2003. Differential SERM effects on corepressor binding dictate ERα activity in vivo. J. Biol. Chem. 278, 6912–6920. Wu, G., Wei, Q., Yu, D. and Shi, F. 2019. Neonatal genistein exposure disrupts ovarian and uterine development in the mouse by inhibiting cellular proliferation. J. Reprod. Dev. 65, 7–17. Xu, B., Lovre, D. and Mauvais-Jarvis, F. 2017. The effect of selective estrogen receptor modulators on type 2 diabetes onset in women: basic and clinical insights. J. Diabetes Complications. 31, 773–779. Xu, Y., Nedungadi, T.P., Zhu, L., Sobhani, N., Irani, B.G., Davis, K.E., Zhang, X., Zou, F., Gent, L.M., Hahner, L.D., Khan, S.A., Elias, C.F., Elmquist, J.K. and Clegg, D.J. 2011. Distinct hypothalamic neurons mediate estrogenic effects on energy homeostasis and reproduction. Cell. Metab. 14, 453–465. You, L., Casanova, M., Bartolucci, E.J., Fryczynski, M.W., Dorman, D.C., Everitt, J.I., Gaido, K.W., Ross, S.M. and Heck, H.d.A. 2002. Combined effects of dietary phytoestrogen and synthetic endocrine-active compound on reproductive development in Sprague-Dawley rats: genistein and methoxychlor. Toxicol. Sci. 66, 91–104. Zhang, J., Song, H., Lu, Y., Chen, H., Jiang, S. and Li, L. 2016. Effects of estradiol on VEGF and bFGF by Akt in endometrial cancer cells are mediated through the NF-κB pathway. Oncol. Rep. 36, 705–714. | ||
| How to Cite this Article |
| Pubmed Style Elsayed D, Helmy S, Dessouki A, El-nahla A, Abdelrazek H, El-hak HG. Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Vet. J.. 2022; 12(5): 639-648. doi:10.5455/OVJ.2022.v12.i5.8 Web Style Elsayed D, Helmy S, Dessouki A, El-nahla A, Abdelrazek H, El-hak HG. Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. https://www.openveterinaryjournal.com/?mno=11804 [Access: June 26, 2026]. doi:10.5455/OVJ.2022.v12.i5.8 AMA (American Medical Association) Style Elsayed D, Helmy S, Dessouki A, El-nahla A, Abdelrazek H, El-hak HG. Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Vet. J.. 2022; 12(5): 639-648. doi:10.5455/OVJ.2022.v12.i5.8 Vancouver/ICMJE Style Elsayed D, Helmy S, Dessouki A, El-nahla A, Abdelrazek H, El-hak HG. Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Vet. J.. (2022), [cited June 26, 2026]; 12(5): 639-648. doi:10.5455/OVJ.2022.v12.i5.8 Harvard Style Elsayed, D., Helmy, . S., Dessouki, . A., El-nahla, . A., Abdelrazek, . H. & El-hak, . H. G. (2022) Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Vet. J., 12 (5), 639-648. doi:10.5455/OVJ.2022.v12.i5.8 Turabian Style Elsayed, Doaa, Seham Helmy, Amina Dessouki, Amal El-nahla, Heba Abdelrazek, and Heba Gad El-hak. 2022. Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Veterinary Journal, 12 (5), 639-648. doi:10.5455/OVJ.2022.v12.i5.8 Chicago Style Elsayed, Doaa, Seham Helmy, Amina Dessouki, Amal El-nahla, Heba Abdelrazek, and Heba Gad El-hak. "Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor." Open Veterinary Journal 12 (2022), 639-648. doi:10.5455/OVJ.2022.v12.i5.8 MLA (The Modern Language Association) Style Elsayed, Doaa, Seham Helmy, Amina Dessouki, Amal El-nahla, Heba Abdelrazek, and Heba Gad El-hak. "Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor." Open Veterinary Journal 12.5 (2022), 639-648. Print. doi:10.5455/OVJ.2022.v12.i5.8 APA (American Psychological Association) Style Elsayed, D., Helmy, . S., Dessouki, . A., El-nahla, . A., Abdelrazek, . H. & El-hak, . H. G. (2022) Influence of genistein and diadizine on regularity of estrous cycle in cyclic female Wistar rat: interaction with estradiol receptors and vascular endothelial growth factor. Open Veterinary Journal, 12 (5), 639-648. doi:10.5455/OVJ.2022.v12.i5.8 |

