




| Original Article | ||
Open Vet. J.. 2023; 13(3): 270-277 Open Veterinary Journal, (2023), Vol. 13(3): 270–277 Original Research Identification of reference genes for expression studies in the liver and spleen of laying hens housed in cage and cage-free systemsMaría P. Herrera-Sánchez1,2, Kelly J. Lozano-Villegas1,2, Iang S. Rondón-Barragán1,2 and Roy Rodríguez-Hernández1*1Poultry Research Group, Laboratory of Immunology and Molecular Biology, Faculty of Veterinary Medicine and Zootechnics, University of Tolima, Ibagué, Colombia 2Immunobiology and Pathogenesis Research Group, Laboratory of Immunology and Molecular Biology, Faculty of Veterinary Medicine and Zootechnics, University of Tolima, Ibagué, Colombia *Corresponding Author: Roy Rodríguez-Hernández. Poultry Research Group, Laboratory of Immunology and Molecular Biology, Faculty of Veterinary Medicine and Zootechnics, University of Tolima, Ibagué, Colombia. Email: royrodriguezh [at] ut.edu.co. Submitted: 23/12/2022 Accepted: 08/02/2023 Published: 05/03/2023 © 2023 Open Veterinary Journal
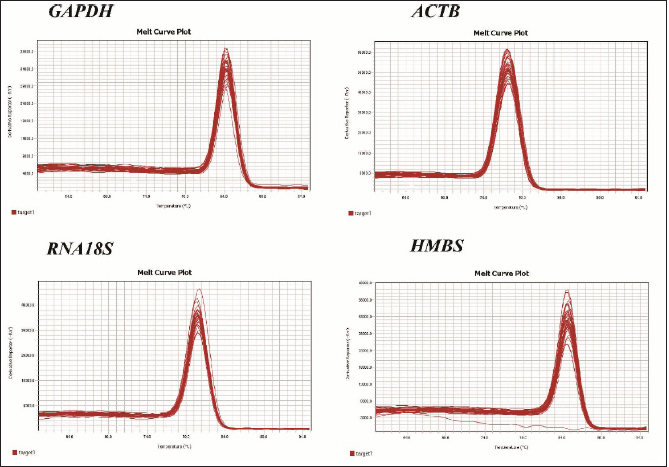
AbstractBackground: The liver and spleen play a pivotal role in metabolism and immune response. During stress, neuroendocrine response induces changes in gene expression, and its assessment demands the validation of the stability of the reference genes to perform relative gene expression experiments. Aim: The objective of this study was to determine the expression stability of four reference genes (GAPDH, ACTB, RNA18S, and HMBS) in the liver and spleen tissues from laying hens housed in a conventional cage (CC) and cage-free (CF) egg production systems. Methods: Liver and spleen from Hy-Line Brown hens housed in CC and CF egg production systems were used. mRNA transcript levels were determined by quantitative polymerase chain reaction (qPCR), and the gene expression stability was evaluated using geNorm, BestKeeper, and NormFinder algorithms. Results: The most stable gene from liver tissue was ACTB in CC, CF, and CC-CF groups (overall data). In the spleen, the most stable genes were GAPDH (CC), HMBS (CF), and ACTB (CC-CF). Conclusion: The ACTB gene was the most stable gene in the liver, and GAPDH and HMBS genes were stable in spleen tissues that could be used for the normalization in qPCR experiments performed in liver and spleen tissues of laying hens housed CC and CF production systems. Keywords: Laying hens, Liver, mRNA, Reference genes, Spleen. IntroductionChicken meat and eggs are the most consumed foods worldwide and continue to be a primary source of quality animal protein in developed and developing countries, contributing to global food security (Mench et al., 2011; Mottet and Tempio, 2017). Currently, the concern about animal welfare, particularly in food production animals, has been growing among consumers (Schuck-Paim et al., 2021). As a result, intensive commercial egg production systems are being evaluated, particularly in conventional cages (CCs), due to the restriction of moving and the inability to perform natural behaviors such as nesting, perching, and dust bathing, generating stress in birds (Løtvedt et al., 2017). Stress factors produce physiological behaviors and effective performance changes and affect gene transcription (Guo et al., 2020; Rostagno, 2020). In the stress responses, activation of the hypothalamic-pituitary-adrenal axis allows the release of glucocorticoids producing a physiologic response that affects the immune function and induces apoptosis of immature spleen and thymus cells, leading to the degeneration of primary lymphoid tissues, including the spleen, thymus, and bursa (Guo et al., 2020). The spleen is the largest immune organ of birds and is primarily responsible for regulating cellular and humoral immunity. However, the neuroendocrine response induced by stress alters the regulation of immune-related genes (El-Lethey et al., 2003; Zhang et al., 2018). Furthermore, in the case of the liver, multiple metabolic functions, including plasma protein synthesis, vitamin, glycogen storage, and fatty acid synthesis, have been reported in birds, which can be affected by glucocorticoids allowing the development of diseases in the long term (Hu et al., 2019). In studies of gene expression, variables such as cDNA concentration and differences in tissues and cells’ gene expression must be controlled (Vandesompele et al., 2002). Therefore, the relative gene expression method needs a reference gene as an internal control whose expression under experimental conditions is unaffected (Vandesompele et al., 2002). Therefore, validating the reference genes’ stability is necessary to consider them adequate for practical needs (Rocha-Martins et al., 2012). This study aimed to determine the expression stability of four reference genes (GAPDH, ACTB, RNA18S, and HMBS) in the liver and spleen tissues of laying hens housed in CC and CF egg production systems. Materials and MethodsStudy populationThe study was carried out using tissue samples from a previous project in the Laboratory of Immunology and Molecular Biology at the University of Tolima. Briefly, under commercial conditions, approximately 60,000 Hy-Line Brown pullets were placed in cages with a density of 16 pullets/cage (314.645 cm2/bird). Pullets were reared with the same sanitary conditions, management, and feed program until 15 weeks of age. Later, they were transferred into two different production systems, conventional cage (CC) and cage-free (CF), on the same farm, up to 82 weeks of age. A total of 45,000 hens were housed in CCs with 4 hens per cage (450 cm2/hen) and 15 replicates of 12 cages each (48 birds/replicate). For the CF system (deep litter), approximately 14,500 hens (1,111 cm2/bird) were distributed in 2 poultry houses, 15 rooms with 990 hens/room. Both systems offered water and the same diet, as well as health and nutritional management following company policies (Rodríguez-Hernández et al., 2021). Samples, RNA extraction, cDNA synthesis, and endpoint polymerase chain reaction (PCR)At 80 weeks of production, six hens (n=6) from CC and six hens (n=6) from CF were randomly selected, and 0.5 g of liver and spleen were extracted. Total RNA was extracted from tissue samples using an RNA-solv reagent kit (OMEGA, Norcross, GA) according to the manufacturer’s instructions. RNA concentration and quality were measured using the NanoDrop One (Thermo Scientific, Wilmington, DE), and cDNA was synthesized with GoScriptTM Reverse Transcription System kit (Promega, Madison, WI) following the manufacturer's instructions. Endpoint PCR and agarose gel electrophoresis of all genes were carried out to determine the cDNA quality and the amplicon size. The reaction had a total volume of 25 µl, composed of 14.8 µl of distilled-deionized water, 5 µl of 5× green GoTaq® Flexi Buffer (Promega, Madison, WI), 1 µl of dNTPs (1.5 mM) (Invitrogen, Carlsbad, CA), 1 µl of each primer (forward and reverse) (10 pmol/µl) (Table 1), 1 µl MgCl2 (25 mM), 0.125 µl of GoTaq® Flexi DNA polymerase (Promega, Madison, WI) and 1 µl of the cDNA as template. The amplifications were carried out in a ProFlex PCR System (Applied Biosystems, Carlsbad, CA) with an initial denaturation step at 95°C for 3 minutes, followed by 35 cycles of denaturation at 95°C for 30 seconds, annealing at 55°C for 30 seconds, extension at 72°C for 30 seconds and a last step of final extension at 72°C for 5 minutes. Amplicons were revealed on 1% agarose gel by electrophoresis (PowerPac™ HC, Bio-Rad, Bio-Rad, Hercules, CA) stained with HydraGreen™ (ACTGene, Piscataway, NJ) and visualized under UV light, using the ENDUROTM GDS gel documentation system (Labnet International, Inc, Woodbridge, NJ). Quantitative PCR (qPCR)Relative gene expression of GAPDH, ACTB, RNA18S, and HMBS genes (Table 1) was measured by qPCR using Luna® Universal qPCR Master Mix (New England BioLabs Inc., Beverly, MA) in a QuantStudio 3 Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA), by fast ramp program. Thermal cycling conditions were initial denaturation of 1 minute at 95°C, then 40 cycles of denaturation for 15 seconds at 95°C, and annealing of 30 seconds at 60°C. Subsequently, a melting step was performed at 95°C for 1 second, 60°C for 20 seconds, and a continuous rise in temperature to 95°C at a rate of 0.15°C per second. Each sample was run in triplicate. Analysis of reference gene expression stabilityExpression levels of the tested reference genes were quantified by the cycle of quantification (Cq) values obtained through qPCR. The three technical replicates were averaged and transformed by the 2−ΔCt method (Zhang et al., 2021). Those values were used as input data on geNorm, and NormFinder, to evaluate the gene expression stability (Vandesompele et al., 2002; Andersen et al., 2004). In the case of the BestKeeper, original data was used (Pfaffl et al., 2004). In addition, analysis of comprehensive data was performed regardless of its origin (CC-CF group). Ethical approvalAll procedures were approved by the Ethics Committee of the University of Tolima, Act 007-2020, based on the Colombia Laws. ResultsPrimer specificityAll the primers used were specific, and a single peak in melt curve analysis indicated no contamination with genomic DNA, primer dimers, or nonspecific PCR products (Fig. 1). Expression profiles of reference genesThe Cq values of the four reference genes from the liver and spleen from laying hens from the CC ranged between 9.30 and 32 and in CF between 9.10 and 27.49. In the CC group, the RNA18S gene is the gene with the highest expression, with Cq values being from 9.30 to 14.10, followed by ACTB (16.13–25.52), GAPDH (16.82–24.88) and HMBS (24.21–32.42) (Fig. 2). On the other hand, in the CF group, as well, the gene most highly expressed was RNA18S (9.10–11.01), followed by ACTB (15.10–19.58), GAPDH (18.34–20.06), and HMBS (24.04–27.49). Table 1. Primers used in the selection of candidate reference genes.
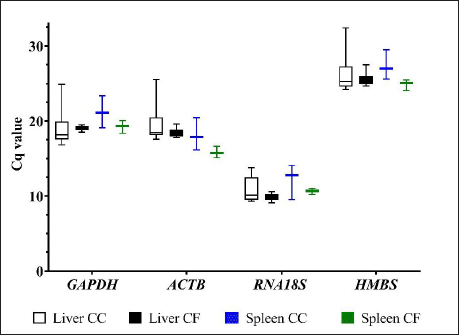
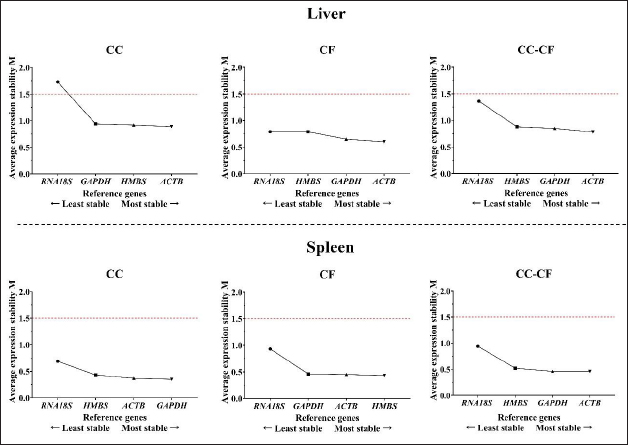
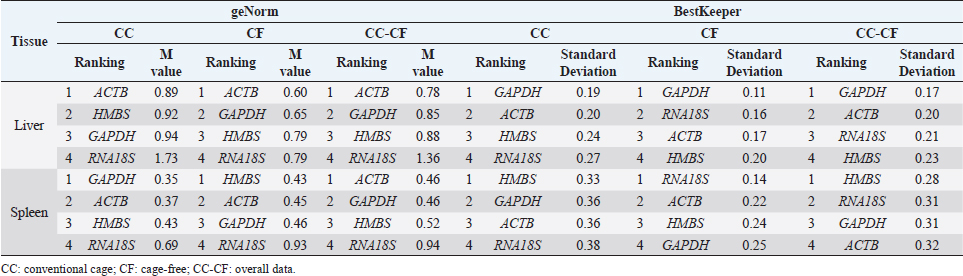
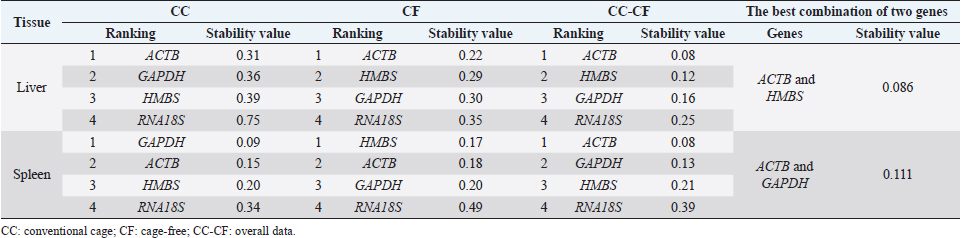
Fig. 1. Melting curve of GAPDH, ACTB, RNA18S, and HMBS genes in the liver and spleen of laying hens. Reference gene stabilityAccording to the geNorm algorithm, a stable reference gene has a M value below 1.5, and in our study, except for the RNA18S gene in the liver (CC) (Fig. 3), all the reference genes were under this value. In the liver, the most stable gene was ACTB, regardless of the production system; and in the spleen, the most stable genes were GAPDH (CC), HMBS (CF), and ACTB (CC-CF) (Table 2). On the other hand, BestKeeper software indicates that the most stable reference gene has a standard deviation (SD) <1, and in the liver, the GAPDH is the most stable gene with an SD of 0.19 (CC), 0.11 (CF), and 0.17 (CC-CF). Furthermore, the genes HMBS (CC and CC-CF) and RNA18S (CF) were stable in the spleen (Table 2). Additionally, NormFinder results showed that ACTB was the stable gene for the liver and spleen in CC-CF. Furthermore, the best combination of genes was ACTB and HMBS for the liver (0.086) and GAPDH and ACTB (0.11) for the spleen (Table 3). DiscussionTranscriptome analysis using technologies such as DNA microarrays, RNA-Seq, and other methods is in demand to evaluate gene expression (Lovén et al., 2012; Hasanpur et al., 2022). However, the qPCR remains a valid and preferred tool to validate the results obtained by transcriptomic analyses (Garrido et al., 2020; Hasanpur et al., 2022). Normalization strategies are needed to achieve accurate results, and reference genes as markers of stability is a common and effective method used for qPCR normalization in the relative quantification method (Vandesompele et al., 2002; Samiullah et al., 2017; Garrido et al., 2020). Nevertheless, the selection and validation of the stability of reference genes for each condition are suggested due to the variation among different experimental conditions and tissues (Wong and Medrano, 2005; Greer et al., 2010; Peng et al., 2018).
Fig. 2.Cq values for four reference genes, GAPDH, ACTB, RNA18S, and HMBS genes in the liver and spleen of laying hens. The Cq values of the GAPDH, ACTB, RNA18S, and HMBS reference genes from liver CC (white boxes), liver CF (black boxes), spleen CC (blue boxes), and spleen CF (green boxes). The box indicates the 25th and 75th percentiles, the lines represent the median, the squares represent the means, and the whiskers represent the maximum and minimum values.
Fig. 3. M value of four candidate reference genes in liver and spleen tissues from laying hens housed in a cage and cage-free systems used the geNorm algorithm. CC: conventional cage; CF: cage-free; CC-CF: overall data. Table 2. Reference gene stability value ranked by geNorm and BestKeeper algorithm in liver and spleen of laying hens of the 80 weeks of production.
Table 3. Reference gene stability value ranked by NormFinder algorithm in liver and spleen of laying hens of the 80 weeks of production.
Several studies have been conducted on screening reference genes in various tissues of laying hens and geese (Nascimento et al., 2015; Zhang et al., 2021). For example, using next-generation sequencing data, Hasanpur et al. (2022) report the Ap2m1 gene as a suitable reference gene for the liver and the Rpl6 gene as a relatively appropriate reference gene for the spleen in chicken. In contrast, Mogilicherla et al. (2022) reported the ALB gene in the liver and the GAPDH gene in the spleen as the most stable reference genes in Gallus gallus. Alternatively, Zhang et al. (2021) identified the ACTB gene as a good reference gene in different tissues of 120-day-old Hy-line brown layer hens. Nevertheless, the reference genes cited previously were defined for other experimental conditions. The geNorm, BestKeeper, and NormFinder algorithms were used to evaluate the stability of the reference genes. The geNorm algorithm calculates the gene expression stability (M) as the mean, the standard deviation of the log-transformed expression ratios for every candidate reference gene under different experimental conditions, where genes with the lowest M represent the most stable gene expression (Vandesompele et al., 2002; Garrido et al., 2020). Regarding NormFinder, this algorithm calculates the most stable gene with the lowest stability value calculated by combining the intragroup and intergroup variations of each gene (Andersen et al., 2004). Finally, the BestKeeper algorithm determines gene expression stability based on the standard deviation and the coefficient of variation (Pfaffl et al., 2004). In this study, the rankings of reference genes were different when using different algorithms. The discrepancy in results delivered by different algorithms has been reported in previous studies (Velada et al., 2015; Gao et al., 2017; Peng et al., 2018). Therefore, the most stable reference genes were selected based on the results from the three algorithms. The liver plays a determining role in the physiological adaptation of animals to changes in the environment, and its gene expression could be affected by different stressors (Kumar et al., 2019; Xu et al., 2019). In the liver, the ACTB gene was classified as the most stable in two of the three algorithms applied. This gene encodes an actin protein involved in cell motility, structure, and integrity (Gonzaga et al., 2020). This result agrees with the findings of Gonzaga et al. (2020), in which the ACTB gene was considered the most stable gene in the liver of two chicken genotypes under heat stress. Furthermore, numerous studies commonly use the ACTB gene as a reference gene (Chapman and Waldenström, 2015). This result differs from the findings of Mogilicherla et al. (2022), where the ALB gene was the most stable in the liver. However, this study’s results suggest that the ACTB gene may not be affected by the egg production system (CC or CF). In addition, using more than one reference gene is recommended to get a more robust, accurate, and reliable normalization of gene expression data (Vandesompele et al., 2002). In this study, the use of GAPDH with the ACTB gene is recommended due to the stable results. In the case of the spleen, it mediates the immune response, and distinct stress factors could alter the regulation of immune response genes (Zhang et al., 2018). The best combination of genes in this tissue was ACTB, GAPDH, and HMBS in all the algorithms applied. Comparable results were demonstrated in chickens under the infectious bronchitis virus infection, where the ACTB gene was suggested as a stable reference gene (Khan et al., 2017). Also, Mogilicherla et al. (2022) described using the GAPDH gene in the spleen for normalization in gene expression studies in chickens. Similarly, Boo et al. (2020) reported that chickens could use GAPDH and HMBS as reference genes. Moreover, in yellow-feathered broilers, the use of the genes ACTB and HMBS were reported as reference genes in different tissues (Zhang et al., 2018). Previously, in magnum tissue of hens housed in CF systems, the most stable gene was HMBS, similar to our results in the spleen of hens of the CF (geNorm and NormFinder) and comprehensive data (CC-CF; BestKeeper) group (Rodríguez-Hernández et al., 2021). On the other hand, the HMBS gene was reported as the most stable gene in the spleen of quail (Macario et al., 2022) and codifying for HMBS protein, a vital enzyme in the heme biosynthetic pathway (Wang et al., 2020). In this study, in the overall data (CC-CF), the ACTB gene was the most stable gene according to two of three software used, which indicate that under the experimental conditions, CC and CF systems do not affect the stability of the gene. Therefore, using the ACTB gene may allow more excellent reliability of qPCR data analysis of hens under the two egg production systems. Furthermore, the best combination of genes was ACTB/HMBS (Liver) and ACTB/GAPDH (Spleen) references genes. AcknowledgmentsWe thank the Laboratory of Immunology and Molecular Biology and the Office of Research-Creation, Innovation, Extension, and Social Projection at the University of Tolima for supporting this research project. Conflicts of interestThe authors declare that they have no conflicts of interest. Author contributionsMaría Herrera-Sánchez and Roy Rodríguez-Hernández were responsible for the design of the study; María Herrera-Sánchez and Kelly Lozano-Villegas performed the experiments; María Herrera-Sánchez analyzed the data; María Herrera-Sánchez and Kelly Lozano-Villegas wrote the manuscript; María Herrera-Sánchez, Kelly Lozano-Villegas, Roy Rodríguez-Hernández, and Iang Rondón-Barragán reviewed and editing the paper. Iang Rondón-Barragán revised the manuscript critically. All authors read and approved the final manuscript. ReferencesAndersen, C.L., Jensen, J.L. and Ørntoft, T.F. 2004. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64(15), 5245–5250. Boo, S.Y., Tan, S.W., Alitheen, N.B., Ho, C.H., Omar, A.R. and Yeap, S.K. 2020. Identification of reference genes in chicken intraepithelial lymphocyte natural killer cells infected with very-virulent infectious bursal disease virus. Sci. Rep. 10(1), 8561. Chapman, J.R. and Waldenström, J. 2015. With reference-to-reference genes: a systematic review of endogenous controls in gene expression studies. PLoS One 10(11), e0141853. El-Lethey, H., Huber-Eicher, B. and Jungi, T.W. 2003. Exploration of stress-induced immunosuppression in chickens reveals both stress-resistant and stress-susceptible antigen responses. Vet. Immunol. Immunopathol. 95(3–4), 91–101. Gao, M., Liu, Y., Ma, X., Shuai, Q., Gai, J. and Li, Y. 2017. Evaluation of reference genes for normalization of gene expression using quantitative RT-PCR under aluminum, cadmium, and heat stresses in soybean. PLoS One 12(1), e0168965. Garrido, J., Aguilar, M. and Prieto, P. 2020. Identification and validation of reference genes for RT-qPCR normalization in wheat meiosis. Sci. Rep. 10(1), 1–12. Greer, S., Honeywell, R., Geletu, M., Arulanandam, R. and Raptis, L. 2010. Housekeeping genes: expression levels may change with the density of cultured cells. J. Immunol. Methods 355(1–2), 76–79. Gonzaga, J., Cedraz de Oliveira, H., Diniz, D.B., Pinto, A.A., Farias, R.V., Gromboni, C.F., Machado, T. and Arias, A. 2020. Influence of heat stress on reference genes stability in heart and liver of two chickens genotypes. PLoS One 15(2), e0228314. Guo, Y., Su, A., Tian, H., Zhai, M., Li, W., Tian, Y., Li, K., Sun, G., Jiang, R., Han, R., Yan, F. and Kang, X. 2020. Transcriptomic analysis of spleen revealed mechanism of dexamethasone-induced immune suppression in chicks. Genes. 11(5), 513. Hasanpur, K., Hosseinzadeh, S., Mirzaaghayi, A. and Alijani, S. 2022. Investigation of chicken housekeeping genes using next-generation sequencing data. Front. Genet. 13, 827538. Hu, X., Wang, Y., Sheikhahmadi, A., Li, X., Buyse, J., Lin, H. and Song, Z. 2019. Effects of glucocorticoids on lipid metabolism and AMPK in broiler chickens' liver. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 232(1), 23–30. Khan, S., Roberts, J. and Wu, S.B. 2017. Reference gene selection for gene expression study in shell gland and spleen of laying hens challenged with infectious bronchitis virus. Sci. Rep. 7(1), 14271. Kumar, H., Iskender, A.U., Srikanth, K., Kim, H., Zhunushov, A.T., Chooq, H., Jang, G., Lim, Y., Song, K.D. and Park, J.E. 2019. Transcriptome of chicken liver tissues reveals the candidate genes and pathways responsible for adaptation into two different climatic conditions. Animals 9(12), 1076. Løtvedt, P., Fallahshahroudi, A., Bektic, L., Altimiras, J. and Jensen, P. 2017. Chicken domestication changes expression of stress-related genes in brain, pituitary and adrenals. Neurobiol. Stress 7, 113–121. Lovén, J., Orlando, D.A., Sigova, A.A., Lin, C.Y., Rahl, P.B., Burge, C.B., Levens, D.L., Lee, T.I. and Young, R.A. 2012. Revisiting global gene expression analysis. Cell 151(3), 476–482. Macario, M.D.S., Nascimento, C.S., Sousa, F.C.B., Oliveira, I.R.S., Vesco, A.P.D., Barbosa, L.T. and Sousa, K.R.S. 2022. Identification of reference genes for studies of quantitative gene expression in male and female quail tissues. Anim. Biotechnol. 6, 1–14. Mench, J.A., Sumner, D.A. and Rosen-Molina, J.T. 2011. Sustainability of egg production in the United States—the policy and market context. Poult. Sci. 90(1), 229–240. Mogilicherla, K., Athe, R., Chatterjee, R. and Bhattacharya, T. 2022. Identification of suitable reference genes for normalization of quantitative real-time PCR-based gene expression in chicken (Gallus gallus). Anim. Genet. 53(6), 881–887. Mottet, A. and Tempio, G. 2017. Global poultry production: current state and future outlook and challenges. Worlds Poult. Sci. J. 73(2), 245–256. Nascimento, C.S., Barbosa, L.T., Brito, C., Roberta, P.M., Mann, R.S., Pinto, A.P., Oliveira, H., Dodson, H., Guimarães, S. and Duarte, M. 2015. Identification of suitable reference genes for real time quantitative polymerase chain reaction assays on pectoralis major muscle in chicken (Gallus gallus). PLoS One 10(5), e0127935. Peng, S., Liu, L., Zhao, H., Wang, H. and Li, H. 2018. Selection and validation of reference genes for quantitative real-time PCR normalization under ethanol stress conditions in Oenococcus oeni SD-2a. Front. Microbiol. 9, 892. Pfaffl, M.W., Tichopad, A., Prgomet, C. and Neuvians, T.P. 2004. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–excel-based tool using pair-wise correlations. Biotechnol. Lett. 26(6), 509–515. Rocha-Martins, M., Njaine, B. and Silveira, M.S. 2012. Avoiding pitfalls of internal controls: validation of reference genes for analysis by qRT-PCR and Western blot throughout rat retinal development. PLoS One 7(8), e43028. Rodríguez-Hernández, R., Oviedo-Rondón, E.O. and Rondón-Barragán, I.S. 2021. Identification of reliable reference genes for expression studies in the magnum of laying hens housed in cage and cage-free systems. Vet. Med. Sci. 7(5), 1890–1898. Rostagno, M.H. 2020. Effects of heat stress on the gut health of poultry. J. Anim. Sci. 98(4), skaa090. Samiullah, S., Roberts, J. and Wu, S.B. 2017. Reference gene selection for the shell gland of laying hens in response to time-points of eggshell formation and nicarbazin. PLoS One 12(7), e0180432. Schuck-Paim, C., Negro-Calduch, E. and Alonso, W.J. 2021. Laying hen mortality in different indoor housing systems: a meta-analysis of data from commercial farms in sixteen countries. Sci. Rep. 11(1), 1–13. Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A. and Speleman, F. 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3(7), 1–12. Velada, I., Ragonezi, C., Arnholdt-Schmitt, B. and Cardoso, H. 2015. Reference genes selection and normalization of oxidative stress responsive genes upon different temperature stress conditions in Hypericum Perforatum L. PLoS One 10(3), e0119982. Wang, Y., Zhang, J., Patrick, K., Li, M., Gong, J., Xu, B., Shen, Q., Yang, Y., Wei, L., Zhang, Y., Peng, D., Ye, J., Poudel, A. and Wang, C. 2020. Hydroxymethylbilane synthase (HMBS) gene-based endogenous internal control for avian species. AMB Express 10(1), 181. Wong, M.L. and Medrano, J.F. 2005. Real-time PCR for mRNA quantitation. BioTech 39(1), 75–85. Xu, E., Zhang, L., Yang, H., Shen, L., Feng, Y., Ren, M. and Xiao, Y. 2019. Transcriptome profiling of the liver among the prenatal and postnatal stages in chickens. Poult. Sci. 98(12), 7030–7040. Zhang, Y., Zhou, Y., Sun, G., Li, K., Li, Z., Su, A., Liu, X., Li, G., Jiang, R., Han, R., Tian, Y., Kang, X. and Yan, F. 2018. Transcriptome profile in bursa of fabricius reveals potential mode for stress-influenced immune function in chicken stress model. BMC Genomics 19(1), 918. Zhang, B.B., Shen, X., Li, X.J., Tian, Y.B., Ouyang, H.J. and Huang, Y.M. 2021. Reference gene selection for expression studies in the reproductive axis tissues of Magang geese at different reproductive stages under light treatment. Sci. Rep. 11(1), 1–10. | ||
| How to Cite this Article |
| Pubmed Style Herrera-sánchez MP, Lozano-villegas KJ, Rondón-barragán IS, Rodríguez-hernández R. Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Vet. J.. 2023; 13(3): 270-277. doi:10.5455/OVJ.2023.v13.i3.3 Web Style Herrera-sánchez MP, Lozano-villegas KJ, Rondón-barragán IS, Rodríguez-hernández R. Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. https://www.openveterinaryjournal.com/?mno=133560 [Access: January 25, 2026]. doi:10.5455/OVJ.2023.v13.i3.3 AMA (American Medical Association) Style Herrera-sánchez MP, Lozano-villegas KJ, Rondón-barragán IS, Rodríguez-hernández R. Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Vet. J.. 2023; 13(3): 270-277. doi:10.5455/OVJ.2023.v13.i3.3 Vancouver/ICMJE Style Herrera-sánchez MP, Lozano-villegas KJ, Rondón-barragán IS, Rodríguez-hernández R. Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Vet. J.. (2023), [cited January 25, 2026]; 13(3): 270-277. doi:10.5455/OVJ.2023.v13.i3.3 Harvard Style Herrera-sánchez, M. P., Lozano-villegas, . K. J., Rondón-barragán, . I. S. & Rodríguez-hernández, . R. (2023) Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Vet. J., 13 (3), 270-277. doi:10.5455/OVJ.2023.v13.i3.3 Turabian Style Herrera-sánchez, María Paula, Kelly Johanna Lozano-villegas, Iang Schroniltgen Rondón-barragán, and Roy Rodríguez-hernández. 2023. Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Veterinary Journal, 13 (3), 270-277. doi:10.5455/OVJ.2023.v13.i3.3 Chicago Style Herrera-sánchez, María Paula, Kelly Johanna Lozano-villegas, Iang Schroniltgen Rondón-barragán, and Roy Rodríguez-hernández. "Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems." Open Veterinary Journal 13 (2023), 270-277. doi:10.5455/OVJ.2023.v13.i3.3 MLA (The Modern Language Association) Style Herrera-sánchez, María Paula, Kelly Johanna Lozano-villegas, Iang Schroniltgen Rondón-barragán, and Roy Rodríguez-hernández. "Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems." Open Veterinary Journal 13.3 (2023), 270-277. Print. doi:10.5455/OVJ.2023.v13.i3.3 APA (American Psychological Association) Style Herrera-sánchez, M. P., Lozano-villegas, . K. J., Rondón-barragán, . I. S. & Rodríguez-hernández, . R. (2023) Identification of reference genes for expression studies in the liver, and spleen of laying hens housed in cage and cage-free systems. Open Veterinary Journal, 13 (3), 270-277. doi:10.5455/OVJ.2023.v13.i3.3 |

