




| Short Communication | ||
Open Vet. J.. 2024; 14(7): 1701-1707 Open Veterinary Journal, (2024), Vol. 14(7): 1701–1707 Short Communication A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR geneHa Thi Thanh Tran†, Anh Duc Truong†, Anh Tuan Tran, Nhu Thi Chu, Vinh The Nguyen, and Hoang Vu Dang**Corresponding Author: Hoang Vu Dang. Department of Biochemistry and Immunology, National Institute of Veterinary Research, Hanoi, Vietnam. Email: dangvuhoang [at] nivr.gov.vn Submitted: 15/04/2024 Accepted: 21/06/2024 Published: 31/07/2024 © 2024 Open Veterinary Journal
AbstractBackground: In 2021, Vietnam experienced an outbreak of Lumpy skin disease (LSD), which infected 207,687 cattle and buffaloes, as officially reported, and resulted in the culling of 29,182 animals. Aim: In this study, samples from cattle that died and showed typical signs of LSD in the Ha Tinh province of Vietnam were confirmed by three World Organization for Animal Health (WOAH)-recommended methods and further studied to compare the Vietnam and China reference strains to the new clinical cases. Methods: Three methods recommended by WOAH for agent detection (PCR, virus isolation, and transmission electron microscopy) were used to confirm this clinical LSD case. The sequence analysis of three well-known markers (P32, RPO30, and GPCR genes) has been utilized in Vietnam to understand this circulating pathogen better. Results: Our findings showed that the CX01 LSDV strain is 100% identical to the Vietnam reference strain HL01 and China reference strains based on P32 and RPO30 genes. Interestingly, analysis of the nucleotide sequence of the GPCR gene showed that the CX01 strain belongs to the same cluster as the reference strains, but it has branches different from those of both the HL01 and China LSDV strains. The nucleotide identification between the CX01 strain and these reference virus strains ranked 99.65%–99.91%, suggesting that it is a new variant of LSDV. Conclusion: This finding is new and indicates that at least two variants of the LSD virus were circulating in Vietnam based on analysis of the GPCR gene. Additionally, these results suggest that the sequence analysis of the GPCR gene is a great tool for subgrouping LSDV circulating in Vietnam. Keywords: Lumpy skin disease, MDBK, Diagnosis, GPCR, Vaccine. BackgroundLumpy skin disease (LSD) represents a resurging viral affliction affecting cattle and buffaloes, resulting in significant economic loss, and its severity prompted the World Organization for Animal Health (WOAH) to classify LSD as a notifiable disease (WOAH, 2021). Transmitted by blood-feeding vectors like mosquitoes and flies, the LSD virus (LSDV) exploits favorable climates to propagate swiftly (Sanz-Bernardo et al., 2021). The LSD epidemic in Europe and Asia has left a profound mark, impacting thousands of livestock and causing substantial financial setbacks. These include losses incurred from animal deaths, diminished productivity, expenses associated with containment efforts, and the forfeiture of export opportunities (Casal et al., 2018). Clinical manifestations of LSD range from subclinical to fatal, contingent upon strain virulence and the susceptibility of cattle breeds. Generally, mortality rates hover below 10%, while morbidity rates can soar up to 90% (OIE, 2017; Sprygin et al., 2018a; WOAH, 2021). Historically confined to southern and eastern Africa since its detection in Zambia in 1929, LSD saw its first outbreak beyond the continent in Egypt and Israel between 1988 and 1989 (Beard, 2016). Today, it maintains an endemic presence in most African nations and has traversed southeastern Europe, the Middle East, and South Asia (Acharya and Subedi, 2020). Recent outbreaks have also been reported in China, India, Bangladesh, and Nepal (Acharya and Subedi, 2020; Lu et al., 2020; Sudhakar et al., 2020; Hasib et al., 2021). The LSD outbreak was first reported in Vietnam in October 2020 (Tran et al., 2020). In addition, over 200,000 cattle and buffaloes were infected, and approximately 30,000 animals were culled during the new Vietnamese outbreak in 2021. These losses were significantly reduced in 2022, as only 2,274 cattle and buffaloes were infected (455 animals culled), indicating success in LSD vaccine strategies (DAH, 2023). The vaccination with the live-attenuated LSDV vaccines was used for the Neethling virus strain, and the goat pox vaccine has been used as a key to the control of the disease in Vietnam. Currently, the Vietnamese Government has approved three live-attenuated LSDV vaccines (ISI-Lympyvac and 2 vaccines using Neethling strain) and one goat pox vaccine, and these vaccines play an important role in the national vaccine program to control and prevent this disease in Vietnam. The homologous of vaccine strains and LSD field strains in Vietnam was more than 99% with the GPCR gene and 100% with the p32 gene. Bioinformatics and phylogenetic tree analysis of LSDV strain using WGS or partial genomes such as GPCR, P32, RPO30, ORF103, or TK genes of LSDV have classified the LSDV into two subgroups (Biswas et al., 2020; Kononov et al., 2019; Ma et al., 2022; Santhamani et al., 2014). The current pool of available nucleotide sequences divides the LSDV into three major phylogenetically distinct lineages: clade 1.1, clade 1.2, and clade 2.5. Clade 1.1 includes classical Neethling-type field strains and is restricted to Africa. Clade 1.2 includes Kenyan/KSGP type field strains that are predominantly present in Africa, the Middle East, Eurasia, and Indian subcontinents (India, Pakistan, Nepal, Srilanka, Bangladesh, Bhutan). The majority of the recombinant LSDV strains are grouped under clade 2.5, which are predominantly circulating in Southeast Asia, including Russia, China, Vietnam, Thailand, and Taiwan (Agianniotaki et al., 2017; Flannery et al., 2022; Ma et al., 2022; Mathijs et al., 2020; Mathijs et al., 2021). The minor populations of clades 2.1, 2.2, 2.3, and 2.4 is present in Russia and Kazakhstan. Previously, it was assumed that these recombinant strains emerged naturally in the field (Kononov et al., 2019). However, later it was confirmed that this was the result of the vaccine spillover and generated due to mishandling of the multiple LSDV strains in the cell culture in the laboratory in Kazakhstan (Vandenbussche et al., 2022), and later, via the vaccine injected into the animals in Kazakhstan. From Kazakhstan, these recombinant vaccine-like strains spread to Russia and China. Nevertheless, natural recombination events in capripoxviruses have never been demonstrated (Vandenbussche et al., 2022). So far, these recombinant strains have never been reported in India. Furthermore, WGS analysis of a novel Saratov/2017 LSDV strain in southern Russia in 2017 and LSDV/Hong Kong/2020 showed similarities with the LSDV Neethling vaccine strain (Sprygin et al., 2018b; Flannery et al., 2022), but the origin of Saratov/2017 and LSDV/Hong Kong/2020 are unclear. Previous studies have indicated that the LSDV strains circulating in Vietnam and China were 100% identical based on three key marker genes (P32, RPO30, and GPCR genes). In this study, samples from cattle that died and showed typical signs of LSD in the Ha Tinh province of Vietnam (2021) were confirmed by three WOAH-recommended methods. In addition, a phylogenetic analysis of three well-known markers was further studied to compare the Vietnam and China reference strains to the new clinical cases. Materials and MethodsSamples collectionThree cattle that died after an outbreak in the Ha Tinh province of Vietnam (2021) were examined based on clinical signs and post-mortem lesions. Blood and tissue samples, including lymph nodes, trachea, spleen, kidney, small intestine, nodules on the skin, mucous membranes, and liver, were collected according to the guidelines of WOAH (WOAH, 2021). The genomic DNA of the capripox virus samples was isolated using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany). The DNA of the capripox virus in the samples was detected by conventional PCR for viral P32 gene-specific primer, and the expected amplicon size was 192 bp (WOAH, 2021). The LSDV Vietnam/Langson/HL01 strain was used as a positive control (Tran et al. 2020), and negative control amplification consisted of nuclease-free sterile water. Virus isolation and titrationVietnam/Langson/HL01 isolated (2020) in the Lang Son province of Vietnam was used as a reference strain (Tran et al. 2020), and the LSDV Vietnam/HaTinh/CX01 strain was collected from the field in Ha Tinh Province of Vietnam. The Department of Biochemistry and Immunology at the Vietnam National Institute of Veterinary Research employed the Madin-Darby bovine kidney (MDBK) cell line to adapt and propagate the virus. These MDBK cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA, USA), supplemented with 100 IU/ml penicillin, 100 mg/ml streptomycin, fungizone (1μg/ml; Sigma-Aldrich; St. Louis, MO, USA), and 5% heat-inactivated fetal bovine serum (FBS; Invitrogen), within a 5% CO2 humidified environment at 37°C. The culture underwent three freeze-thaw cycles at −80°C, followed by thorough mixing, and the resulting clarified supernatant was subsequently introduced to fresh cultures for successive passages until characteristic cytopathogenic effects (CPE) were observed under an inverted microscope. The presence of CPE and fluctuations in viral titers across passages confirmed the permissive nature of the cell culture for the virus. Titration of the LSDV was conducted in 96-well tissue culture plates, utilising the Reed and Muench method for calculation, with results expressed in log TCID50/ml (Reed and Muench, 1938). The stock titers of LSDV strains ranged from 106.5 to 107.5 TCID50/ml after five passages on the MDBK cell line. Transmission electron microscopySeventy-two hours post-infection with LSDV, MDBK cells were harvested from the culture flask using a cell scraper followed by centrifugation at 3,000 × g for 5 minutes. The cells were then suspended, fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.2), and left overnight at 4°C. Subsequently, the cells underwent four careful rinses in 0.1 M cacodylate buffer (pH 7.2). The final cell pellet was fixed in 1% OsO4 in the same buffer for 60 minutes, followed by another four rinses in 0.1 M cacodylate buffer (pH 7.2). The cells were then dehydrated in a series of ethanol solutions (ranging from 50% to 100%), washed in propylene oxide, and immersed for 6 hours in a 1:1 mixture of propylene oxide and epoxydic resin for infiltration. Following embedding in the resin, thin sections were obtained using an ultramicrotome (UltracutUC6; Leica Microsystems GmbH, Germany) and stained with uranyl acetate and lead citrate at the National Institute of Hygiene and Epidemiology in Hanoi, Vietnam. Finally, the ultrathin sections were examined using a transmission electron microscope operating at 80 kV. Genomic characterization of LSDV strainThe DNA extraction from the samples was conducted for PCR amplification. The correct size of the amplicons of the partial gene P32, GPCR, and RPO30 products were electrophoresed on a 1.5% agarose gel against a 100 bp DNA leader marker (Thermo Scientific) and visualized by UV irradiation and ethidium bromide staining (Sigma-Aldrich, St. Louis, MS, USA) (Santhamani et al., 2014; El-Tholoth and El-Kenawy, 2016; Tran et al., 2020). Amplicons of the correct size were excised from the agarose gel and purified using the QIAQuick gel extraction kit (QIAgen) according to the manufacturer’s specifications for sequencing (1st BASE, Selangor, Malaysia). The chromatograms of amplicon sequences were analyzed using BioEdit and the DNAstar program (DNASTAR Inc. Madison, WI, USA). The nucleotide identity of the LSDV strain in Vietnam was compared with other sequences using the Blast tool at the National Center for Biotechnology Information (NCBI) database and using the information of published sequences. The Lasergene software (DNASTAR Inc.) performed multiple sequence alignment. Phylogenetic analyses of nucleotide sequences of partial or complete coding sequences of P32, GPCR, and RPO30 gene LSDV were constructed using the neighbor-joining method with a bootstrap value of 1,000 in the MEGA7 program (Kumar et al., 2016). Ethical approvalThe study complied with the institutional rules for the care and use of laboratory animals and a protocol approved by the Ministry of Agriculture and Rural Development, Vietnam (TCVN 8402:2010). Results and DiscussionFrom January to December 2021, a total of 13,463 family farms in 1,330 villages (208 communes of 13 districts) in the Ha Tinh Province of Vietnam officially reported an LSD outbreak with a total of 17,611 infected cattle and buffaloes. In total, 2,934 animals were culled during this period, indicating severe losses to the cattle industry in this province (DAH, 2023). The clinical signs and pathological lesions of castles are critical for the diagnosis of LSDV in the field (OIE, 2017; WOAH, 2021). As shown in Figure 1, cattle from the Ha Tinh province of Vietnam that died and exhibited typical clinical signs, including nodules on the skin, mucous membranes and internal organs, emaciation, enlarged lymph nodes, and oedema of the skin, were identified. Importantly, the nodules in the skin were found in the entire body of cattle and the trachea of dead cattle. Furthermore, the small intestine and mesenteric lymph nodes were hemorrhagic and turned a blackish colour. According to the WOAH, nodules are mainly present in the body and internal organs, and the lymph nodes are hemorrhagic in all cases (WOAH, 2021). Additionally, the malnourished animal could potentially suffer from pneumonia and mastitis. Additionally, it may exhibit necrotic skin plugs, possibly resulting from fly infestation, leading to the formation of deep holes in the hide. These same findings suggested the presence of LSDV in the cattle mortality from the Ha Tinh province of Vietnam.
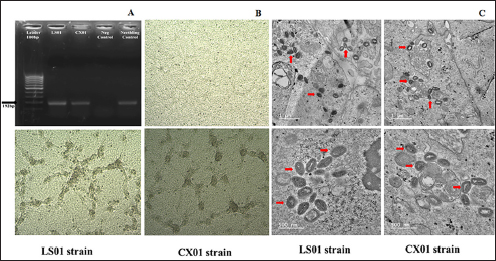
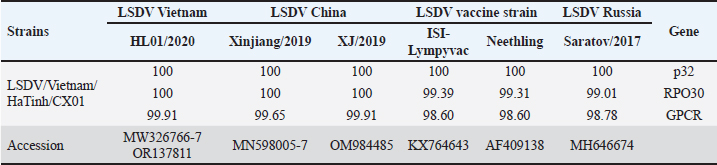
Fig. 1. Clinical signs of lumpy skin disease in cattle in the study area in Ha Tinh province of Vietnam. Nodule lesion in mouth (A), Nodule lesion in back leg (B), Nodule lesion in font leg (C), hemorrhage in small intestine (D), nodule lesion in trachea (E), and hemorrhage in intestinal lymph node (F). To confirm the presence of LSDV in the clinical case, three WOAH-recommended methods for agent detection were performed, including PCR, virus isolation, and transmission electron microscopy. First, DNA extracted from skin nodules was used as a template for the conventional PCR to confirm the presence of the DNA of LSDV, and the results demonstrated that the samples were positive for LSD (Fig. 2A). LSDV from the clinical samples were isolated and propagated in MDBK cells lines. The characteristic CPE in the MDBK cell line is shown in Figure (2B. This effect was marked by cell rounding and clustering, forming rosette-shaped aggregations scattered across the monolayer sheet after 24 hours post-infection (hpi). Over time, these aggregations increased, ultimately resulting in the detachment of the cell monolayer after 96 hpi. By this point, most sheets exhibited vacuoles due to cell detachment, with the remaining cells either aggregated or elongated. Notably, the CPE was observed solely in the infected cells, not the uninfected control cells, as depicted in Figure 2B. This observation confirmed the presence of LSDV in the field sample. Furthermore, electron microscopy imaging revealed a membrane surrounding the capsid structure (Fig. 2C). Viral particles were identified within the MDBK cell cultures. These virions exhibited a large, mature, ovoid shape with rounded ends. Notably, the viral particles presented as ovoid electron-dense bodies, displaying the characteristic short mulberry-like features, as shown in Figure 2C. According to the WOAH, PCR, virus isolation, and electron microscopy methods are recommended for LSDV diagnosis from field samples (WOAH, 2021). These results also confirmed the success of LSDV isolation from the clinical samples. This strain is named as LSDV/Vietnam/HaTinh/CX01. For genetic analysis of the CX01 LSDV strain, PCR sequencing was used to analyze the genetic information of CX01 LSDV compared to the LSDV/Vietnam/Langson/HL01 strain (a well-known reference strain from the first outbreaks in Vietnam). Three primer sets were used to amplify the P32 gene (LSDV074), RP030 (LSDV036) gene, and GPCR (LSDV011) gene, as previously described (Zhu et al., 2013; OIE, 2017; Sameea Yousefi and al., 2017; Alexander et al., 2019; Kononov et al., 2019; Ochwo et al., 2020;). The results indicated that the nucleotide comparison between the LSDV/Vietnam/HaTinh/CX01 virus strain and the LSDV/Vietnam/Langson/HL01 (MW326769), as well as the China strain Xinjiang/2019 (MN598005) and the China/XJ/2019 (MN518933), was 100% identical based on the P32 and RP030 (Table 1, Fig. 3B and C). Additionally, it was 100% and 99.39% identical for the ISI-lumpyvax strain, 100% and 99.31% identical for the Neethling strain, and 100% and 90.01% identical for Saratov/2017, as shown in Table 1. In contrast, using the BLAST tool from NCBI to identify the nucleotide of GPCR of CX01 LSDV strain with other reference virus strains, it was 99.91% identical with the HL01 strain (GenBank accession no. OR137811), 99.65% identical with the Xinjiang/2019 strain, 98.60% identical with two vaccine strains, and 98.87% identical with Saratov/2017 strain (Table 1). A recent study indicated that in the GPCR gene, the nucleotide sequence mutation or amino acid mutation, such as addition or deletion, was found in some LSDV strain field samples. The resulting GPCR gene is suitable for use in the host-range grouping of capripoxviruses and subgrouping of LSDV (Santhamani et al., 2014; El-Tholoth and El-Kenawy, 2016; Cetre-Sossah et al., 2017). Furthermore, the GPCR genes have played an important role in the viral ligand-to-cell receptors, cell activation, replication, virulence, neutralizing antibody response, or responses to antiviral drugs (Le Goff et al., 2009). Research in Russia (2017), based on nucleotide and amino acid sequence of the GPCR gene protein, the Saratov/2017 LSDV strain was identified and classified into a new vaccine-like LSDV strain circulating in Russian cattle (Kononov et al., 2019). The findings suggest that the emergence of new variants within LSDV strains is a significant concern. This is particularly concerning due to the potential for the circulation of both field and vaccine strains, it may be contributed to the development of a new variant of LSDV (Fig. 3A). Interestingly, the LSDV/Vietnam/HaTinh/CX01 strain displays a 12-bp insertion homologous to the vaccine strain. Our findings reveal that the field strains are segregated into new variants based on GPCR analysis. These findings would be helpful to Vietnam in considering any concerns in the development of a vaccine program using the field strain to develop inactivated, live attenuated LSDV or recombinant vaccine programs to control prevalent LSDV in Vietnam.
Fig. 2. LSDV Confirmation in clinical case by three WOAH recommended methods (A) Gel photograph of p32 gene PCR (B) LSDV strains LS01 and CX01 with cytopathic effect (CPE) at the second passage on MDBK cell line (below) and uninfected MDBK cell line after 96 hpi and (C) Transmission electron micrograph showing large LSDV particles in the MDBK of infected cells as a brick shape with rounded ends and characteristic short surface tubules (mulberry) with Scale bar: 500 nm and 1μm. Table 1. The nucleotide identity of LSDV/Vietnam/HaTinh/CX01 strain in north-central Vietnam in comparison with LSDV/Vietnam/Langson/HL01 isolated in the first outbreaks in Vietnam and other reference sequences was performed using the Blast tool at the NCBI database and using the information of published sequence.
Fig. 3. Phylogenetic tree based on GPCR gene (A), P32 gene (B), and RPO30 gene (C) sequences showing the genetic relationship of LSDV strains from Ha Tinh province in north-central Vietnam with LSDV reference strains. The sequences obtained from this study are labeled in red triangles. Other LSDV, SPPV, and GTPV sequences analyzed are from GenBank. Bootstrap values of 1,000 replicates are shown when higher than 70%. The consensus tree was rooted in reference to the three poxvirus outgroups and was broken up to show the distant relationship between them and CaPVs. The collective findings underscore the importance of conducting a comprehensive analysis of the entire LSDV genome to monitor potential alterations and the further spread of the virus. Controlling the transmission of the LSDV is crucial, alongside investigating its prevalence and determining the most appropriate vaccination strategy in Vietnam. Additionally, various measures, such as enhancing biosafety awareness, intensifying oversight of illegal animal movements and product transport, and reducing the population density of LSDV vectors, could mitigate the risk of transmission within Vietnam. Finally, the development of a safe and effective LSDV vaccine stands as the optimal long-term solution to safeguard Vietnam and other nations against the virus. ConclusionWe demonstrated that the LSDV/Vietnam/HaTinh/CX01 strain isolate belongs to a new variant of LSDV, and the GPCR gene's sequencing analysis is of great concern for sub-grouping LSDV circulating in Vietnam. This finding is new and suggests that at least two different variants of LSDV are circulating in Vietnam based on analysis of the GPCR gene. Additionally, the combination methods, including PCR, virus isolation, and electronic electron microscopy imaging, were powerful methods to confirm the presence of LSDV in clinical samples. AcknowledgmentsThe authors thank the staff of Region Animal Health Office 3 in Nghe An for their support in sampling. The author also thanks Dr. Andrew G. Yersin of Kemin Industries, Inc., Scott Avenue, Des Moines, IA 50317, USA, for the English revision. Conflict of interestThere are no potential conflicts of interest. FundingThe Ministry of Science and Technology supported this work to Ha Thi Thanh Tran (Project code: ĐTĐL.CN-11/23). Data availabilityAll sequences of LSDV/Vietnam/HaTinh/CX01 strain generated in this study were submitted to GenBank under Accession Nos OR137808-10 for p32 genes, RPO30 genes, GPCR, and genes, respectively. Authors’ contributionsHTTT, ADT, and HVD conceived and designed the experiments. HTTT, ADT, ATT, CTN, and VNT performed the experiments. HTTT, ADT, and HVD analyzed the data. HVD and HTTT contributed reagents, materials, and analytical tools. HTTT, ADT, and HVD wrote the manuscript. All authors read and approved the final manuscript. ReferencesAcharya, K.P., Subedi, D., 2020. First outbreak of lumpy skin disease in Nepal. Transbound. Emerg. Dis. 67(6), 2280–2281. Agianniotaki, E.I., Mathijs, E., Vandenbussche, F., Tasioudi, K.E., Haegeman, A., Iliadou, P., Chaintoutis, S.C., Dovas, C.I., Van Borm, S., Chondrokouki, E.D., De Clercq, K., 2017. Complete genome sequence of the lumpy skin disease virus isolated from the first reported case in Greece in 2015. Genome Announc. 5(29), e00550–17. Alexander, S., Olga, B., Svetlana, K., Valeriy, Z., Yana, P., Pavel, P., Aleksandr, K., 2019. A real-time PCR screening assay for the universal detection of lumpy skin disease virus DNA. BMC Res. Notes 12, 371. Beard, P.M., 2016. Lumpy skin disease: a direct threat to Europe. Vet. Rec. 178, 557–558. Biswas, S., Noyce, R.S., Babiuk, L.A., Lung, O., Bulach, D.M., Bowden, T.R., Boyle, D.B., Babiuk, S., Evans, D.H., 2020. Extended sequencing of vaccine and wild-type capripoxvirus isolates provides insights into genes modulating virulence and host range. Transbound. Emerg. Dis. 67, 80–97. Casal, J., Allepuz, A., Miteva, A., Pite, L., Tabakovsky, B., Terzievski, D., Alexandrov, T. and Beltran-Alcrudo, D. 2018. Economic cost of lumpy skin disease outbreaks in three Balkan countries: Albania, Bulgaria and the Former Yugoslav Republic of Macedonia (2016-2017). Transbound. Emerg. Dis. 65(6), 1680–1688. Cetre-Sossah, C., Dickmu, S., Kwiatek, O., Albina, E., 2017. A G-protein-coupled chemokine receptor: A putative insertion site for a multi-pathogen recombinant capripoxvirus vaccine strategy. J. Immunol. Methods 448, 112–115. Departmen Animal Health (DAH), Vietnam., 2023. Anumal repport of diseases in animal, Vietnam, 2023. Offically Anumal Report. DAH, Hanoi, Vietnam. El-Tholoth, M., El-Kenawy, A.A., 2016. G-protein-coupled chemokine receptor gene in lumpy skin disease virus isolates from cattle and water buffalo (Bubalus bubalis) in Egypt. Transbound. Emerg. Dis. 63, e288–e295. Flannery, J., Shih, B., Haga, I.R., Ashby, M., Corla, A., King, S., Freimanis, G., Polo, N., Tse, A.C., Brackman, C.J., Chan, J., Pun, P., Ferguson, A.D., Law, A., Lycett, S., Batten, C., Beard, P.M., 2022. A novel strain of lumpy skin disease virus causes clinical disease in cattle in Hong Kong. Transbound. Emerg. Dis. 69, e336–e343. Hasib, F.M.Y., Islam, M.S., Das, T., Rana, E.A., Uddin, M.H., Bayzid, M., Nath, C., Hossain, M.A., Masuduzzaman, M., Das, S., Alim, M.A., 2021. Lumpy skin disease outbreak in cattle population of Chattogram, Bangladesh. Vet. Med. Sci. 7, 1616–1624. Kononov, A., Byadovskaya, O., Kononova, S., Yashin, R., Zinyakov, N., Mischenko, V., Perevozchikova, N., Sprygin, A., 2019. Detection of vaccine-like strains of lumpy skin disease virus in outbreaks in Russia in 2017. Arch. Virol. 164, 1575–1585. Kumar, S., Stecher, G., Tamura, K., 2016. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874. Le Goff, C., Lamien, C.E., Fakhfakh, E., Chadeyras, A., Aba-Adulugba, E., Libeau, G., Tuppurainen, E., Wallace, D.B., Adam, T., Silber, R., Gulyaz, V., Madani, H., Caufour, P., Hammami, S., Diallo, A., Albina, E., 2009. Capripoxvirus G-protein-coupled chemokine receptor: a host-range gene suitable for virus animal origin discrimination. J. Gen. Virol. 90, 1967–1977. Lu, G., Xie, J., Luo, J., Shao, R., Jia, K., Li, S., 2020. Lumpy skin disease outbreaks in China, since 3 August 2019. Transbound. Emerg. Dis. 68(2), 216–219. Ma, J., Yuan, Y., Shao, J., Sun, M., He, W., Chen, J., Liu, Q., 2022. Genomic characterization of lumpy skin disease virus in southern China. Transbound. Emerg. Dis. 69, 2788–2799. Mathijs, E., Vandenbussche, F., Ivanova, E., Haegeman, A., Aerts, L., De Leeuw, I., Van Borm, S., De Clercq, K., 2020. Complete coding sequence of a lumpy skin disease virus from an outbreak in Bulgaria in 2016. Microbiol. Resour. Announc. 9(43), e00977–20. Mathijs, E., Vandenbussche, F., Nguyen, L., Aerts, L., Nguyen, T., De Leeuw, I., Quang, M., Nguyen, H.D., Philips, W., Dam, T.V., Haegeman, A., Van Borm, S., De Clercq, K., 2021. Coding-complete sequences of recombinant lumpy skin disease viruses collected in 2020 from four outbreaks in Northern Vietnam. Microbiol. Resour. Announc. 10, e0089721. Ochwo, S., VanderWaal, K., Ndekezi, C., Nkamwesiga, J., Munsey, A., Witto, S.G., Nantima, N., Mayanja, F., Okurut, A.R.A., Atuhaire, D.K., Mwiine, F.N., 2020. Molecular detection and phylogenetic analysis of lumpy skin disease virus from outbreaks in Uganda 2017-2018. BMC Vet. Res. 16, 66. OIE, 2017. Chapter 2.4.13. Lumpy skin disease OIE Terrestrial Manual 2017. OIE, Paris, France. Reed, L.J., Muench, H., 1938. A simple method of estimaing fifty percent endpoints. Am. J. Epidemiol. 27, 493–497. Sameea Yousefi, P., Mardani, K., Dalir-Naghadeh, B., Jalilzadeh-Amin, G., 2017. Epidemiological study of lumpy skin disease outbreaks in North-western Iran. Transbound. Emerg. Dis. 64, 1782–1789. Santhamani, R., Yogisharadhya, R., Venkatesan, G., Shivachandra, S.B., Pandey, A.B., Ramakrishnan, M.A., 2014. Molecular characterization of Indian sheeppox and goatpox viruses based on RPO30 and GPCR genes. Virus Genes 49, 286–291. Sanz-Bernardo, B., Suckoo, R., Haga, I.R., Wijesiriwardana, N., Harvey, A., Basu, S., Larner, W., Rooney, S., Sy, V., Langlands, Z., Denison, E., Sanders, C., Atkinson, J., Batten, C., Alphey, L., Darpel, K.E., Gubbins, S. and Beard, P.M. 2021. The Acquisition and Retention of Lumpy Skin Disease Virus by Blood-Feeding Insects Is Influenced by the Source of Virus, the Insect Body Part, and the Time since Feeding. J. Virol. 96(15), e0075122. Sprygin, A., Artyuchova, E., Babin, Y., Prutnikov, P., Kostrova, E., Byadovskaya, O., Kononov, A., 2018a. Epidemiological characterization of lumpy skin disease outbreaks in Russia in 2016. Transbound. Emerg. Dis. 65, 1514–1521. Sprygin, A., Pestova, Y., Prutnikov, P., Kononov, A., 2018b. Detection of vaccine-like lumpy skin disease virus in cattle and Musca domestica L. flies in an outbreak of lumpy skin disease in Russia in 2017. Transbound. Emerg. Dis. 65, 1137–1144. Sudhakar, S.B., Mishra, N., Kalaiyarasu, S., Jhade, S.K., Hemadri, D., Sood, R., Bal, G.C., Nayak, M.K., Pradhan, S.K., Singh, V.P., 2020. Lumpy skin disease (LSD) outbreaks in cattle in Odisha state, India in August 2019: epidemiological features and molecular studies. Transbound. Emerg. Dis. 67(6), 2408–2422. Tran, H.T.T., Truong, A.D., Dang, A.K., Ly, D.V., Nguyen, C.T., Chu, N.T., Hoang, T.V., Nguyen, H.T., Nguyen, V.T., Dang, H.V., 2021. Lumpy skin disease outbreaks in vietnam, 2020. Transbound. Emerg. Dis. 68, 977–980. Vandenbussche, F., Mathijs, E., Philips, W., Saduakassova, M., De Leeuw, I., Sultanov, A., Haegeman, A., De Clercq, K., 2022. Recombinant LSDV strains in Asia: vaccine spillover or natural emergence? Viruses 14(7), 1429. WOAH, 2021. Infection with lumpy skin disease virus. OIE Terrestrial Manual 2021 Chapter 11.9. WOAH, Paris, France. Zhu, X.L., Yang, F., Li, H.X., Dou, Y.X., Meng, X.L., Li, H., Luo, X.N., Cai, X.P., 2013. Identification and phylogenetic analysis of a sheep pox virus isolated from the Ningxia Hui Autonomous Region of China. Genet. Mol. Res. 12, 1670–1678. | ||

| How to Cite this Article |
| Pubmed Style Tran HTT, Truong AD, Tran AT, Chu NT, Nguyen VT, Dang HV. A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Vet. J.. 2024; 14(7): 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 Web Style Tran HTT, Truong AD, Tran AT, Chu NT, Nguyen VT, Dang HV. A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. https://www.openveterinaryjournal.com/?mno=197208 [Access: January 24, 2026]. doi:10.5455/OVJ.2024.v14.i7.19 AMA (American Medical Association) Style Tran HTT, Truong AD, Tran AT, Chu NT, Nguyen VT, Dang HV. A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Vet. J.. 2024; 14(7): 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 Vancouver/ICMJE Style Tran HTT, Truong AD, Tran AT, Chu NT, Nguyen VT, Dang HV. A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Vet. J.. (2024), [cited January 24, 2026]; 14(7): 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 Harvard Style Tran, H. T. T., Truong, . A. D., Tran, . A. T., Chu, . N. T., Nguyen, . V. T. & Dang, . H. V. (2024) A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Vet. J., 14 (7), 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 Turabian Style Tran, Ha Thi Thanh, Anh Duc Truong, Anh Tuan Tran, Nhu Thi Chu, Vinh The Nguyen, and Hoang Vu Dang. 2024. A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Veterinary Journal, 14 (7), 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 Chicago Style Tran, Ha Thi Thanh, Anh Duc Truong, Anh Tuan Tran, Nhu Thi Chu, Vinh The Nguyen, and Hoang Vu Dang. "A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene." Open Veterinary Journal 14 (2024), 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 MLA (The Modern Language Association) Style Tran, Ha Thi Thanh, Anh Duc Truong, Anh Tuan Tran, Nhu Thi Chu, Vinh The Nguyen, and Hoang Vu Dang. "A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene." Open Veterinary Journal 14.7 (2024), 1701-1707. Print. doi:10.5455/OVJ.2024.v14.i7.19 APA (American Psychological Association) Style Tran, H. T. T., Truong, . A. D., Tran, . A. T., Chu, . N. T., Nguyen, . V. T. & Dang, . H. V. (2024) A new variant of lumpy skin disease virus circulating in Vietnam based on sequencing analysis of GPCR gene. Open Veterinary Journal, 14 (7), 1701-1707. doi:10.5455/OVJ.2024.v14.i7.19 |

