




| Research Article | ||
Open Vet. J.. 2024; 14(7): 1689-1700 Open Veterinary Journal, (2024), Vol. 14(7): 1689–1700 Research Article Molecular identification, virulence, and antibiotic-resistant genes characteristics of Staphylococcus spp., isolated from milk samplesAfrah Sabeeh Mhyson1, Masar Saab Kadhim2, Orooba Meteab Faja2*, Merriam Ghadhanfar Alwan3 and Basima Jasim Mohammed2*Corresponding Author: Orooba Meteab Faja. College of Veterinary Medicine, University of Al-Qadisiyah, Al-Diwaniyah City, Iraq. Email: orooba.faja [at] qu.edu.iq Submitted: 21/04/2024 Accepted: 26/06/2024 Published: 31/07/2024 © 2024 Open Veterinary Journal
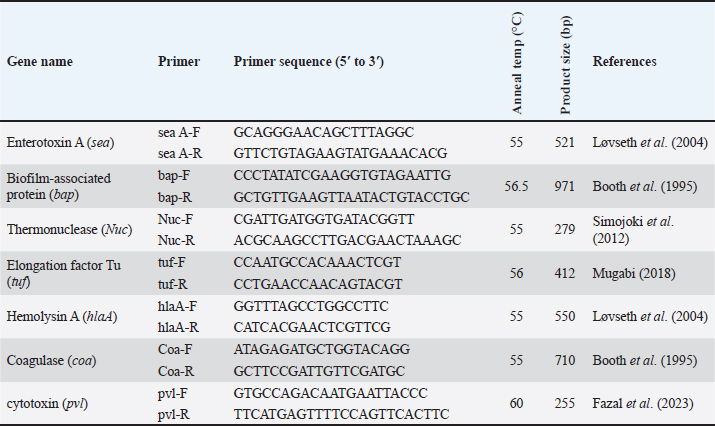
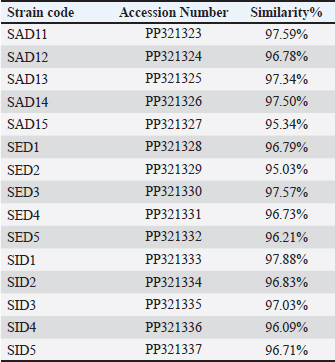
AbstractBackground: Food safety and food-borne infections are major subjects of global interest. Dairy products are considered as important source for these infections. Aim: The present study was conducted to identify the occurrence and to genotype isolates of Staphylococcus spp. recovered from milk samples in Al-Diwaniyah City, Iraq. Methods: The current study included the collection of 50 milk samples purchased from local stores in the current city. These samples were subjected to bacterial cultivation and biochemical tests. Later, the growth was used to extract the genomic DNA that was exposed to PCR and partial gene sequencing both targeted the 16S rRNA gene at a specific genetic piece. Results: The PCR results demonstrated the amplification of the genetic fragment of five genetic clusters for each of Staphylococcus aureus (SAD11, SAD12, SAD13, SAD14, and SAD15), Staphylococcus epidermidis (SED1, SED2, SED3, SED4, and SED5), and Staphylococcus intermedius (SID1, SID2, SID3, SID4, and SID5). The PCR products were sent out to sequencing and reported that the current isolates were similar in their genetic content with global isolates at 95.34% to 97.59%, 96.21% to 97.57%, and 96.09% to 97.88%, respectively, of identity. Conclusion: The present findings show high genetic variations among isolates of S. aureus, S. epidermidis, and S. intermedius recovered from milk samples, and these genotypes are found in different infection settings related to humans and animals, which may pose high risks to humans and animals. Keywords: Food hygiene, Food-borne diseases, Food safety. IntroductionStaphylococcus aureus is a Gram-positive, coagulase-positive, spherical pathogenic bacterial species belonging to the family Staphylococcaceae. It is a commensal inhabitant on many parts of the human body and when present without symptoms it is found on the skin, especially of the anterior nostrils, perineum, groin, and axilla (Lakhundi and Zhang, 2018). Due to its adaptability to hosts and environment, it causes many infections of varying severity, ranging from mild to lobar pneumonia and septicemia. Staphylococcus aureus infections are also considered a major cause of both hospital and community-originated infections. The organism induces disease involving the circulation, cutaneous infection, pneumonia, and foodborne diseases, as well as cause infections by medical devices, such as endocarditis (Diekema et al., 2001; Lindsay and Holden, 2004; Schito, 2006). The bacterium secretes unique illness-inducing virulence factors, such as enterotoxins causing diseases, such as foodborne diseases. Staphylococcus aureus is in a group of bacteria that can build up resistance to a wide range of antimicrobial drugs. The methicillin utilization resulted in the methicillin-resistant S. aureus (MRSA) occurrence, which significantly increased morbidity and mortality. The occurrence of MRSA infections is being increasingly documented globally. There are different MRSA strains, but those commonly seen produce a modified penicillin-binding protein, PBP (2a or 2′) (Lakhundi and Zhang, 2018). This altered PBP has a weak binding affinity to penicillin that is mediated by a 34-amino-acid segment located near the primary binding site. The altered PBP is encoded by the mecA gene, which is clustered on a mec (SCCmec), a chromosomal element (Katayama et al., 2000). Recently, mecC, a less common carrier, was detected in isolates from MRSA, which are resistant most likely to all penicillins, except for some medicines such as ceftobiprole. Moreover, Vancomycin is routinely used mostly because it is more effective in vitro than penicillin. Vancomycin is weighed as the last resort for some severe MRSA infections. However, due to its escalated use, moderate to highly resistant isolates to vancomycin have emerged in countries around the globe. Early identification of MRSA infections, using phenotypic and molecular techniques, and rapid sending of these results to the clinician, could enhance patient care. This might reduce the use of inappropriate antimicrobials, antimicrobial resistance, and cost (Buonomini et al., 2020). Since these bacteria have a wide range of distribution and sources in different environments, such as milk, the present study was conducted to identify the occurrence and genotype isolates of Staphylococcus spp. recovered from milk samples in Al-Diwaniyah City, Iraq. Materials and MethodsSamplesHere, 50 samples of raw milk from different local outlet stores in Al-Diwaniyah City in Iraq have been collected, filled in sterile bottles, and then immediately delivered to the microbiology laboratory in the College of Veterinary Medicine, University of Al-Qadisiyah, in the same city and then stored at refrigerator at 4°C until culturing bacteria. Bacterial cultivationThese specimens were put in Brain Heart Infusion Broth overnight at 37°C for first enrichment, then inoculated into mannitol salt agar, and incubated at 37°C overnight the next day for optimization as to culture just Staphylococcus spp. Molecular identificationFive isolates of each species (the most virulent isolates) were selected for molecular identification by PCR. DNA extractionThe protocol and its Wizard Genomic DNA Purification Kit (Promega) were employed for the extraction of the DNA. From the bacterial broth, 1 ml was used as starting material for the extraction. A NanoDrop was recruited to measure the DNA according to its quality and quantity. PCRThe PCR targeted the 16S rRNA gene by utilizing the sets of the primer; F: AGAGTTTGATCATGGCTCAG and R: GGTTACCTTGTTACGACTT. The reaction mixture contains 20 mM Tris-HCl (pH 8.4), 50 mM KCl, 1.5 mM MgCl2, 2 μl of any of dNTPs, 0.4 μM for the respective primer and 2.5 U DNA polymerase in a volume of 50 μl. The addition of about 10 to 100 ng DNA to the reaction mixture gives to total volume of 50 μl. The PCR was performed using an early denaturation step of 2 minutes at 94°C then 35 cycles of 1 minute at 94°C, 1.5 minutes at 55°C and 1 minute at 72°C, of main denaturation, annealing, and main extension steps, respectively, which finally followed by a final extension step at 72°C for 3 minutes. The expected PCR amplification product has a size of 1500 base pairs. The agarose gel at 1% mixed with ethidium bromide was employed at 80–90 Volt for 60 minutess. Table 1. Primers of the PCR for the virulence gene detection of Staphylococcus spp. isolated from milk samples.
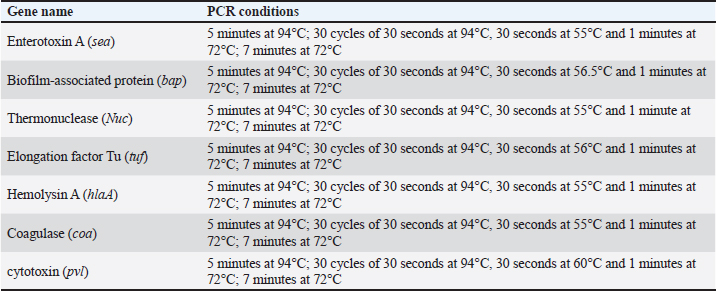
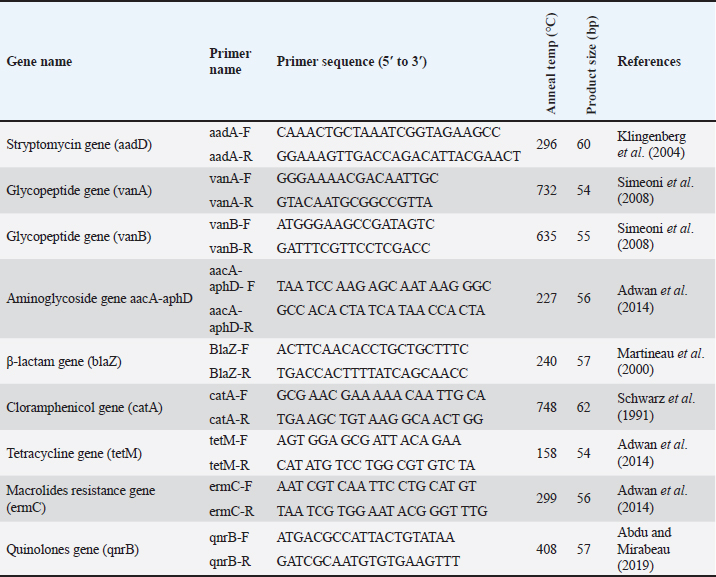
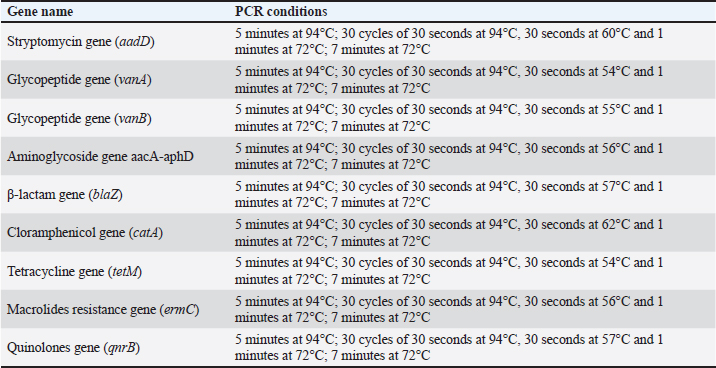
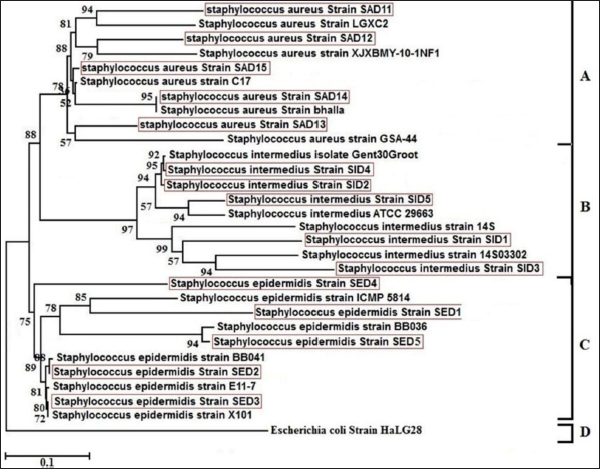
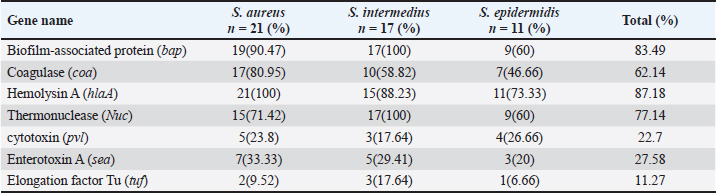
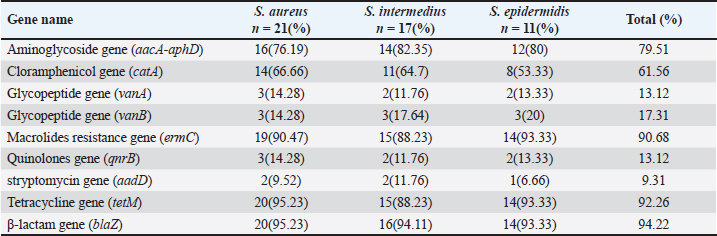
Sequencing of 16S rRNA gene (partial)The PCR-amplified DNA sequences were sent for sequencing by ABI3730XL system (Macrogen Inc. Korea), and phylogenic analysis and tree were performed using NCBI-Websites and MEGA X software. Detection of virulence genesAll isolated bacteria (68 isolates=32 isolates from human specimen and 36 isolates from chicken samples) were screened for the presence of virulence genes. Polymerase chain reaction (PCR) assays were applied to target the virulence determinants enterotoxin gene (stn), host recognition and invasion gene (invA), changes in the interaction of host immune system gene (spvC), fimbrial proteins gene (fimA), iron acquisition gene (iroB), and survival within macrophage gene (msgA) (Chaudhary et al., 2015; Borges et al., 2017; Soubeiga et al., 2022). The details of the primers are displayed in Table 1. Before the experiment, we performed the trial run for each primer for optimization MgCl and annealing temperature, which were displayed in Table 2. 25 μl Vol of reaction was made by PCR master mix (a 2x EconoTaq® PULS GREEN 2X Master Mix, DA5110, Lucigen). Detection of antibiotic resistance genesStreptomycin (aadA1), aminoglycoside (aac(3)11a), β-lactam (blaTEM), Cloramphenicol (catA), Sulfonamides (sul1), Tetracycline (tetA), Macrolides resistance (ermB), and Quinolones (qnrB) were employed (Chuanchuen et al., 2008) and listed in Table 3. For each primer, the PCR conditions are shown in Table 4. Ethical approvalNot needed for this study. ResultsBacterial isolation and identificationOut of 60 milk samples, 53 positive samples (81.66%) were detected. Including 21 S. aureus, 17 Staphylococcus intermedius, and 15 Staphylococcus epidermidis. The PCR results demonstrated the amplification of the genetic fragment of five genetic clusters for each of S. aureus (SAD11, SAD12, SAD13, SAD14, and SAD15), S. epidermidis (SED1, SED2, SED3, SED4, and SED5), and S. intermedius (SID1, SID2, SID3, SID4, and SID5) (Fig. 1). The PCR products were sent out to sequencing and reported that the current isolates were similar in their genetic content with global isolates at 95.34% to 97.59%, 96.21% to 97.57%, and 96.09% to 97.88%, respectively, of identity (Table 5 and Fig. 2). Antibiotics have been developed to treat S. aureus infections, leading to the selection of more virulent strains with mutations that confer resistance to antibiotics. In recent decades, many S. aureus strains have acquired resistance to multiple antibiotics, posing a serious problem in modern clinical practice. As a result, the genotyping of strains has become an urgent task to guide epidemiological study and clinical practice. The present study recognized the following genotypes; SAD12, SAD14, SAD15, SED1, SED2, SED4, SID4, SID5, and SID1. The percentage of virulence genes (bap, coa, hlaA, nuc, pvl, sea, and tuf) of Staphylococcus spp. from this study are presented in Table 6. The detection of virulence genes in Staphylococcus spp. isolates by PCR technique is shown in Figure 3. Result analysis of total Staphylococcus spp. isolates that used in this test showed that hlaA gene has the highest percentage (87.18%) followed by bap, nuc, and coa gene which had 77.14% and 62.14%. The least virulent gene from all tested Staphylococcus spp. isolates were sea, pvl, and tuf genes which had 27.58%, 22.7%, and 11.27%. Under S. aureus isolates category, the highest virulent gene was hlaA which had 100%, followed by bap which had 90.47%, coa with 80.95%, tlh with 37.5%, and the nuc gene was found to be 71.42% and the least under this category was found to be of sea, pvl and tuf genes which had 33.33%, 23.8%, and 9.52%, respectively. Under S. intermedius isolates category, the gene with the highest percentage were bap and nuc genes which were present in all tested isolates (100%), followed by hlaA, coa, and sea which found to be (88.23%, 58.82%, and 29.41%), respectively. The least under this category were tuf and pvl genes with 17.64%. Under S. epidermidis isolates category, the highest virulent gene was hlaA which had 73.33%, followed by nuc and bap, which both had 60%, the intermediate percentages were of coa and pvl with 46.66% and 26.66%, respectively. The least under this category was tuf genes with 6.66%. Table 2. PCR therocycler conditions for the detection of virulance genes of Staphylococcus spp. isolated from milk samples.
Table 3. Primers of the PCR for the antibiotic resistance gene detection of Staphylococcus spp. isolated from milk samples.
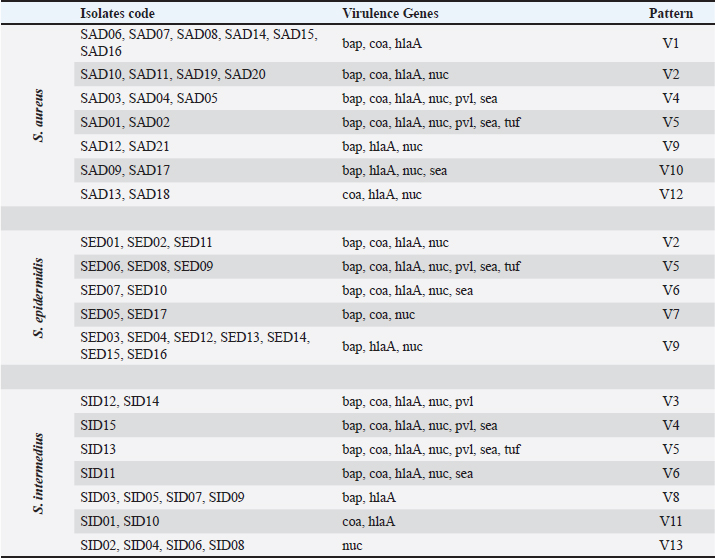
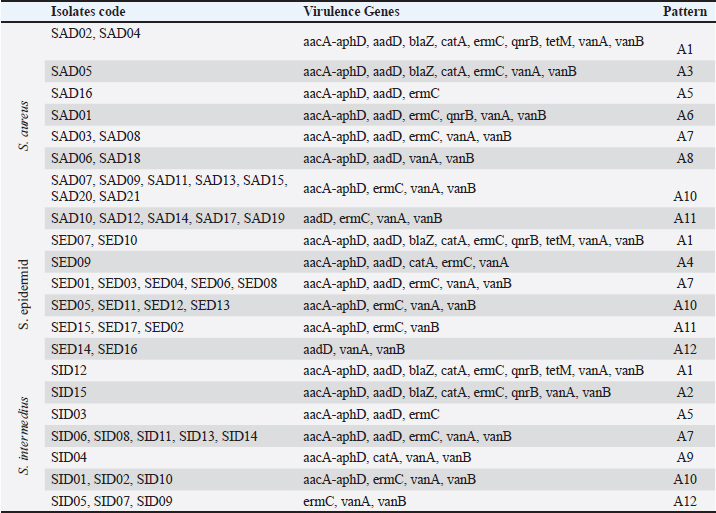
The virulence gene patterns of Staphylococcus spp. isolated in this study is presented in Table 7. Staphylococcus spp. isolates have shown 13 different virulence gene patterns. There were 6 isolates (SAD01, SAD02, SED06, SED08, SED09, and SID13) carrying all tested genes, and the tested isolates were carrying at least one of seven tested virulence genes. The most frequently repeated pattern was V9 (bap, hlaA, nuc) which appeared in nine isolates. The percentages of nine resistance genes of 53 Staphylococcus spp. isolates (21 S. aureus, 17 S. intermedius, and 15 S. epidermidis) are shown in Table 8. The detection of antibiotic resistance genes in Staphylococcus spp. isolates by PCR technique is shown in Figure 4. The data analysis of the total tested isolate showed a variety of percentages from high, moderate to low. The highest percentage was of blaZ with 94.22% followed by tetM and ermCgenes which had 92.26% and 90.68%, respectively. The moderate percentages of resistant genes were of aacA-aphD and catA which had 79.51% and 61.56%, respectively. The lowest percentage of total tested isolates was of vanB, qnrB, and vanA gene which had 17.31%, 13.12%, and 13.12%, respectively. Closer inspection of S. aureus isolates showed that the highest percentage was of blaZ and tetM gene which both had 95.23%, followed by ermC, aacA-aphD, and catA genes which had 90.47%, 76.19%, and 66.66%, respectively. qnrB, vanB and vanA genes were having the same percentage (14.28%). The lowest antibiotic resistance gene percentage of S. aureus isolates was of aadD gene with 9.52%. Table 4. PCR therocycler conditions for the detection ofantibiotic resistance genes of Staphylococcus spp. isolated from milk samples.
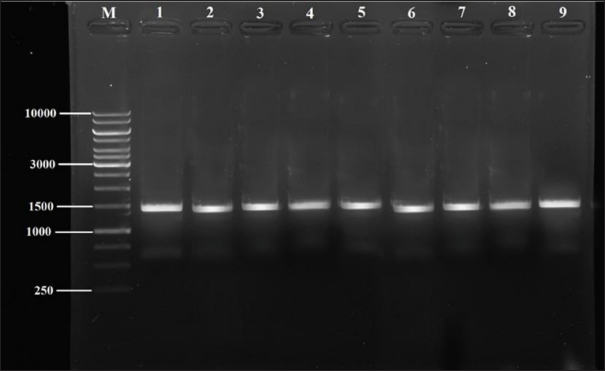
Fig. 1. Image of the 16S rRNA related amplification on 1% agarose gel of nine isolates of Staphylococcus spp from milk samples. M: DNA ladder and Lanes of 1 to 9: SAD12, SAD14, SAD15, SED1, SED2, SED4, SID4, SID5, SID1. Table 5. Identity rates of current study of 16S rRNA related sequencing alignment of nine isolates of Staphylococcus spp from milk samples.
Under the category of S. intermedius isolates, there was no gene with 100% and the highest percentage was of blaZ gene which had 94.11% and followed by tetM, ermC, aacA-aphD and catA genes with 88.23%, 88.23%, 82.35%, and 64.7%, respectively. It is apparent that the lowest percentage of seawater, sediment, and total tested isolates was of aadD, qnrB, and vanA gene, which had 11.76%. Under the category of S. epidermidis isolates, the highest percentage was of blaZ, tetM, and ermC gene which had 93.33% and followed by aacA-aphD, catA, and vanB genes with 80%, 53.33%, and 20%, respectively. The lowest percentage of seawater, sediment, and total tested isolates was of aadD gene, which had 6.66%. Table 9 shows the antibiotic resistance gene patterns of Staphylococcus spp. isolated from seawater and sediment samples. Seven gens were investigated and Staphylococcus spp. isolates have shown 12 different antibiotic resistance gene patterns. The presence of antibiotic-resistance genes was varied from three to nine genes out of nine tested genes. There were five isolates (SAD02, SAD04, SED07, SED10, and SID12) carrying all tested genes and belong to patterns A1 pattern. The most frequently repeated pattern was A10 pattern (aacA-aphD, ermC, vanA, vanB) which was reported in 14 isolates, followed by A7 pattern (aacA-aphD, aadD, ermC, vanA, vanB) which were reported in 12 isolates. All pattern was reported in 8 isolates, A12 and A1 patterns were reported in five isolates each. The other patterns were repeated in lower frequency in tested Staphylococcus spp. isolates.
Fig. 2. Phylogenetic tree of of 16S rRNA related sequencing alignment of nine isolates of Staphylococcus spp from milk samples. Table 6. Percentage of virulence genesof Staphylococcus spp. isolated from milk samples.
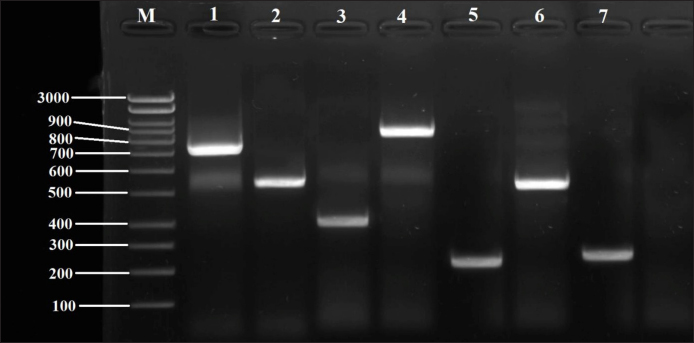
Fig. 3. Detection of virulence genes in Staphylococcus spp. isolates by PCR technique, electrophoresed on 1.5% (w/v) agarose gel. Lane M: 100bp DNA ladder; Lane 1: coa gene in SAD01; Lane 2: hlaA gene in SED06; Lane 3: tuf gene in SED09; Lane 4: bap gene in SAD02; Lane 5: pvl gene in SID13; Lane 6: sea gene in SID15; Lane 7: nuc gene in SAD04. DiscussionFor SAD14 and SAD15 Genotype: A study by Loewen et al. (2017) reports that these two genotypes, SAD14 and SAD15 are two very similar types of S. aureus strains and that these two genotypes seemed to have a relatively heightened occurrence in community-acquired infections. The research also suggests that in the case of SAD14 and SAD15, a relatively lower antibiotic resistance profile could be observed in contrast to SAD12. The researchers concluded that a potential shift in the epidemiology of S. aureus infections could be observed where these two genotypes could gain importance in a community-acquired setting. They argue that the SED1, SED2, and SED4 strains have been demonstrating a greater likelihood to persist long-term, which would imply that they are extremely successful inhabitants, able to thrive in the human host for a long time. Recently it has also been shown that SED4 could play a role in increased antibiotic resistance by finding a high prevalence among emerging antibiotic-resistant strains, of which MRSA was the most popular. In light of this, individual genotyping has been proposed as a potential tool to accurately predict the susceptibility of strains to antibiotics. For SID4, SID5, and SID1 genotype: It was found that the genetic structure of S. aureus establishes the existence of specific genetic DNA markers that set them apart from other genotypes. Specifically, they studied the SID4, SID5, and SID1 genotypes involving well-differentiated strains. SID4 was found to have a relatively higher prevalence in HAIs compared to other genotypes. On the other hand, SID5 and SID1 were reported to be relatively more common among CAIs than other genotypes. Similarly, the research proposes that these three genotypes exhibited specific differences in their antibiotic resistance profiles (Elsayed and Dawoud, 2015). Table 7. Virulence genes patternof Staphylococcus spp. isolated from milk samples.
Table 8. Prevalence of antibiotic resistance genesof Staphylococcus spp. isolated from milk samples.
This result is consistent with what has been found in a previous study of Khasapane et al. (2024) how reported coa gene (42%), and hla gene (38%). The presence of hlaA gene at 38% in Staphylococcus spp. isolates in the present study are in contrast with previous findings in Egypt (34.4% and 43.75%) (Singh et al., 2014). Brazil (38% and 58%) (Mühlberg et al., 2020), and China (57% and 36%). However, this is lower than those from various studies in China, in which the results of hlaA genes were more than 80% (Wang et al., 2016).
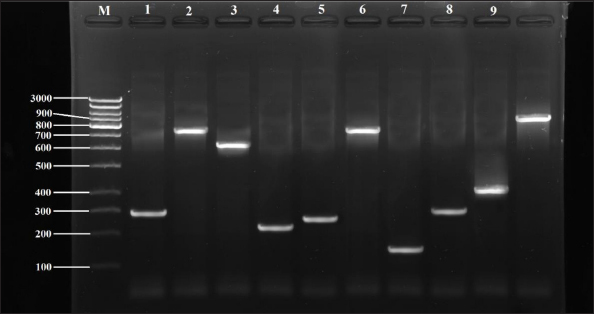
Fig. 4. Detection of virulence genes in Staphylococcus spp. isolates by PCR technique, electrophoresed on 1.5% (w/v) agarose gel. Lane M: 1K DNA ladder; Lane 1: aadD gene in SAD01; Lane 2: vanA gene in SAD01; Lane 3: vanB gene in SED09; Lane 4: aacA-aphD gene in SAD02; Lane 5: blaZ gene in SED10; Lane 6: catA gene in SID15; Lane 7: tetM gene in SAD04; Lane 8: ermC gene in SID15; Lane 9: qnrB gene in SAD04. Table 9. Antibiotic resistance gene patternsof Staphylococcus spp. isolated from milk samples.
In the present study, the identification of the staphylococcal enterotoxin gene sea in the isolates studied was recognized in 27.58% of them. These findings are consistent with previous results obtained by several other authors: German authors (7.10%), Brazilians (10.9%), and Czechs (19.4%). The percentage of isolates that carried the sea gene was significantly lower compared with those obtained in raw meats from the South African market (35.29%), from the market of northern Egypt (52%), and from the supermarkets in Italy (65.60%). The results of pvl gene also were aligned with the previous report of Mello et al. (2016) who reported that 10.7% of staphylococcal carried pvl gene. Previous study of Karimzadeh and Ghassab (2022), they found nuc gene present in 30% of tested Staphylococcus isolates which is interestingly opposite to our finding where nuc gene shown in 77.14%. The coa gene results were in agreement with the previous study of Chmagh and Abd Al-Abbas (2019) who found 77.3% of the tested isolates were carrying coa gene. Salina et al. (2020) was in agreement with this study, where they found that bap gene in Staphylococcus were present in 58% of tested isolates. The result showed that 90.68% of the isolates of Staphylococcus spp. carrying the erm(C) gene while 9.32% were negative for erm(C) (Table 5). These findings was in contrast to a previous study which reveals that the prevalence of erm(A) gene has been established as the predominant dissemination of the methicillin-resistant strains (Schafer and Phillippi, 2020) but conforms with previous studies which reveals the wide distribution and acceptance of the erm(C) in S. aureus isolates. Also, Denmark has an increase occurrence of staphylococcus spp. carrying erm (C) (Qu et al., 2010). Some previous studies have revealed that the prevalence of the erm (A) is more than erm (C) gene in the tested isolates. Current study revealed that most of the isolates of Staphylococcus spp. were positive for tetM; in fact 92.26% of the isolates carried tetM, in contrast to previous studies which revealed that the prevalence of erm (A) gene has been established as the predominant dissemination of the methicillin-resistant strains (Fujiwara et al., 2017; Schafer and Phillippi, 2020) but a previous study by Bagcigil and his colleagues in Turkey conforms with the results of the current work, revealing a high prevalence of tetM in isolates of S. aureus isolates obtained from clinical patients and staphylococcus spp. was revealed to be the leading cause of bovine mastitis (Bagcigil et al., 2012). An opposite result was obtained in the research by Aubry et al. (2020), where 30% of strains were found to contain the blaZ gene (Aubry et al., 2020). Similar to our study, As of Bagcigil et al. (2012) the presence of blaZ gene was determined in S. aureus isolates, determined by broth microdilution, originating from bovine mastitis, and all isolates were found to be positive for the blaZ gene. Other study by Salina et al. (2020) showed seventeen (59%) tested isolates contained the cat gene which were in agreement with our results. Amin et al. (2021) investigated the presence of qnrB gene, they found 6% of Staphylococcus spp were carrying the gene, this result is in consistent with our results where qnrB gene showed 13.12%. in earlier study (Benites et al., 2021), the presence of the aacA-aphD gene, which codes for enzymes that act on aminoglycosides, was detected in 28.1% of the Staphylococcus spp. isolates, this result contradicts our results that shown higher aacA-aphD gene percentage (79.51%). Monecke and Ehricht (2005) investigate the percentages of aadD gene and they found that 29% of Staphylococcus spp. isolates were carrying aadD gene, these findings are not directly in line with our results (9.31%). This result ties well with previous studies (Monecke and Ehricht, 2005) whereinvanA and vanB resistant genes were detected in 34% and 37% of clinical isolates, respectively. One of the main findings in this study was the wide spread of antimicrobial resistance genes among Staphylococcus spp. Resistance genes of blaZ, ermC, and tetM were prevalent. This makes sense as these antibiotics have evolved naturally in the environment and so are primed to select for the spread of the resistance determinants or for selection of locally increasing isolates (that is, resistance evolves due to persistent contact with the antibiotic in the environment, which can increase its use among bacterial populations already present (Rosser and Young, 1999). ConclusionThe present findings show highly genetic variations among isolates of S. aureus, S. epidermidis, and S. intermedius recovered from milk samples, and these genotypes are found in different infection setting related to humans and animals, which may pose high risks to humans and animals. AcknowledgmentsThe authors would like to thank their colleges and institutions. Conflict of interestThe authors declare that there is no conflict of interest for the current study. Authors’ contributionAll authors had participated in samples collection, performing tests, analyzing the data, and preparing and revising the manuscript. FundingThe authors had no specific funding sources for this study but personal funding only. Data availabiltyThe data for the current study are available upon request. ReferencesAbdu, A.B. and Mirabeau, T.Y. 2019. Prevalence of qnr genes among multidrug resistance Staphylococcus aureus from clinical isolates. J. Adv. Med. Med. Res. 30(10), 1–10. Adwan, K., Jarrar, N., Abu-Hijleh, A., Adwan, G. and Awwad, E. 2014. Molecular characterization of Escherichia coli isolates from patients with urinary tract infections in Palestine. J. Med. Microbiol. 63(Pt 2), 229–234. Aubry, B., Lemarié, C., Chenouard, R., Kempf, M., Eveillard, M. and Pailhoriès, H. 2020. Performance of penicillinase detection tests in Staphylococcus epidermidis: Comparison of different phenotypic methods. BMC Microbiol. 20, 1–8. Bagcigil, A.F., Taponen, S., Koort, J., Bengtsson, B., Myllyniemi, A.L. and Pyörälä, S. 2012. Genetic basis of penicillin resistance of S. aureus isolated in bovine mastitis. Acta Vet. Scand. 54, 1–7. Benites, N.R., Hora, A.S.D., Mello, P.L., Laes, M.A., Brandão, P.E., Melville, P.A. and Zuniga, E. 2021. Genotypic evaluation of antimicrobial resistance in Staphylococcus spp. isolated from bovine clinical mastitis. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. 73, 302–310. Booth, M.C., Atkuri, R.V., Nanda, S.K., Iandolo, J.J. and Gilmore, M.S. 1995. Accessory gene regulator controls Staphylococcus aureus virulence in endophthalmitis. Invest. Ophthalmol. Vis. Sci. 36(9), 1828–1836. Borges, I., Faveri, M., Figueiredo, L.C., Duarte, P.M., Retamal-Valdes, B., Montenegro, S.C.L. and Feres, M. 2017. Different antibiotic protocols in the treatment of severe chronic periodontitis: A 1-year randomized trial. J. Clin. Periodontol. 44(8), 822–832. Buonomini, A.R., Riva, E., Di Bonaventura, G. and Gherardi, G. 2020. Rapid detection of methicillin-resistant Staphylococcus aureus directly from blood for the diagnosis of bloodstream infections: a mini-review. Diagnostics. 10, 830. Chaudhary, J.H., Nayak, J.B., Brahmbhatt, M.N. and Makwana, P.P. 2015. Virulence genes detection of Salmonella serovars isolated from pork and slaughterhouse environment in Ahmedabad, Gujarat. Vet. World 8(1), 121–124. Chmagh, A.A. and Abd Al-Abbas, M.J. 2019. Comparison between the coagulase (coa and vwb) genes in Staphylococcus aureus and other staphylococci. Gene Rep. 16, 100410. Chuanchuen, R., Wannaprasat, W., Ajariyakhajorn, K. and Schweizer, H.P. 2008. Role of the MexXY multidrug efflux pump in moderate aminoglycoside resistance in Pseudomonas aeruginosa isolates from Pseudomonas mastitis. Microbiol. Immunol. 52(8), 392–398. Diekema, D.J., Pfaller, M.A., Schmitz, F.J., Smayevsky, J., Bell, J., Jones, R.N., Beach, M. and Group, S.P. 2001. Survey of infections due to Staphylococcus species: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe, and the Western Pacific region for the SENTRY Antimicrobial Surveillance Program, 1997–1999. Clin. Infect. Dis. 32, S114–S132. Elsayed, M.S. and Dawoud, M.A. 2015. Phenotypic and genotypic detection of virulence factors of Staphylococcus aureus isolated from clinical and subclinical mastitis in cattle and water buffaloes from different farms of Sadat City in Egypt. Vet. World 8(9), 1051. Fazal, M.A., Rana, E.A., Akter, S., Alim, M.A., Barua, H. and Ahad, A. 2023. Molecular identification, antimicrobial resistance and virulence gene profiling of Staphylococcus spp. associated with bovine sub-clinical mastitis in Bangladesh. Vet. Anim. Sci. 21, 100297. Fujiwara, N., Tsuruda, K., Iwamoto, Y., Kato, F., Odaki, T., Yamane, N. and Noguchi, M. 2017. Significant increase of oral bacteria in the early pregnancy period in Japanese women. J. Investig. Clin. Dent. 8(1), e12189. Karimzadeh, R. and Ghassab, R.K. 2022. Identification of nuc nuclease and sea enterotoxin genes in Staphylococcus aureus isolates from nasal mucosa of burn hospital staff: a cross-sectional study. New Microbes New infect. 47, 100992. Katayama, Y., Ito, T., Hiramatsu, K. 2000. A new class of genetic element, Staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 44, 1549–1555. Khasapane, N.G., Nkhebenyane, J., Mnisi, Z., Kwenda, S. and Thekisoe, O. 2024. Comprehensive whole genome analysis of Staphylococcus aureus isolates from dairy cows with subclinical mastitis. Front. Microbiol. 15, 1376620. Klingenberg, C., Sundsfjord, A., Rønnestad, A., Mikalsen, J., Gaustad, P. and Flægstad, T. 2004. Phenotypic and genotypic aminoglycoside resistance in blood culture isolates of coagulase-negative staphylococci from a single neonatal intensive care unit, 1989–2000. J. Antimicrob. Chemother. 54(5), 889–896. Lakhundi, S., Zhang, K. 2018. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbiol. Rev. 31, e00020-18. Lindsay, J.A., Holden M.T. 2004. Staphylococcus aureus: Superbug, super genome? Trends Microbiol. 12, 378–385. Loewen, K., Schreiber, Y., Kirlew, M., Bocking, N. and Kelly, L. 2017. Community-associated methicillin-resistant Staphylococcus aureus infection: Literature review and clinical update. Can. Fam. Physician. 63(7), 512–520. Løvseth, A., Loncarevic, S. and Berdal, K.G. 2004. Modified multiplex PCR method for detection of pyrogenic exotoxin genes in staphylococcal isolates. J. Clin. Microbiol. 42(8), 3869–3872. Martineau, F., Picard, F.J., Grenier, L., Roy, P.H., Ouellette, M. and Bergeron, M.G. 2000. Multiplex PCR assays for the detection of clinically relevant antibiotic resistance genes in staphylococci isolated from patients infected after cardiac surgery. J. Antimicrob. Chemother. 46(4), 527–534. Mello, P.L., Moraes Riboli D.F., Pinheiro L., de Almeida Martins L., Vasconcelos Paiva Brito M.A., Ribeiro de Souza da Cunha M.D.L. 2016. Detection of enterotoxigenic potential and determination of clonal profile in Staphylococcus aureus and coagulase-negative staphylococci isolated from bovine subclinical mastitis in different Brazilian states. Toxins. 8(4), 104. Monecke, S. and Ehricht, R. 2005. Rapid genotyping of methicillin-resistant Staphylococcus aureus (MRSA) isolates using miniaturised oligonucleotide arrays. Clin. Microbiol. Infect. 11(10), 825–833. Mugabi, R. 2018. Genotypes and phenotypes of Staphylococci on selected dairy farms in vermont. The University of Vermont and State Agricultural College. Ph.D. Thesis: Available via https://scholarworks.uvm.edu/graddis/844 Mühlberg, E., Umstätter, F., Kleist, C., Domhan, C., Mier, W. and Uhl, P. 2020. Renaissance of vancomycin: approaches for breaking antibiotic resistance in multidrug-resistant bacteria. Can. J. Microbiol. 66(1), 11–16. Qu, Y., Daley, A.J., Istivan, T.S., Rouch, D.A. and Deighton, M.A. 2010. Densely adherent growth mode, rather than extracellular polymer substance matrix build-up ability, contributes to high resistance of Staphylococcus epidermidis biofilms to antibiotics. J. Antimicrob. Chemother. 65(7), 1405–1411. Rosser, S.J. and Young, H.K. 1999. Identification and characterization of class 1 integrons in bacteria from an aquatic environment. J. Antimicrob. Chemother. 44(1), 11–18. Salina, A., Guimaraes, F.F., Pereira, V.B., Menozzi, B.D., Rall, V.L.M. and Langoni, H. 2020. Detection of icaA, icaD, and bap genes and biofilm production in Staphylococcus aureus and non-aureus staphylococci isolated from subclinical and clinical bovine mastitis. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. 72, 1034–1038. Schafer, R. and Phillippi, J.C. 2020. Group B streptococcal bacteriuria in pregnancy: an evidence-based, patient-centered approach to care. J. Midwifery Womens Health. 65(3), 376–381. Schito, G.C. 2006. The importance of the development of antibiotic resistance in Staphylococcus aureus. Clin. Microbiol. Infect. 12, S3–S8. Schwarz, S. and Noble, W.C. 1994. Structure and putative origin of a plasmid from Staphylococcus hyicus that mediates chloramphenicol and streptomycin resistance. Lett. Appl. Microbiol. 18(5), 281–284. Simeoni, D., Rizzotti, L., Cocconcelli, P., Gazzola, S., Dellaglio, F. and Torriani, S. 2008. Antibiotic resistance genes and identification of staphylococci collected from the production chain of swine meat commodities. Food Microbiol. 25(1), 196–201. Simojoki, H., Hyvönen, P., Ferrer, C.P., Taponen, S. and Pyörälä, S. 2012. Is the biofilm formation and slime producing ability of coagulase-negative staphylococci associated with the persistence and severity of intramammary infection? Vet. Microbiol. 158(3-4), 344–352. Singh, M., Singh, A. and Sharma, A. 2014. Production and applications of an N-terminally-truncated recombinant beta-haemolysin from Staphylococcus aureus. Biologicals. 42(4), 191–198. Soubeiga, A.P., Kpoda, D.S., Compaoré, M.K.A., Somda-Belemlougri, A., Kaseko, N., Rouamba, S.S., Ouedraogo, S., Traoré, R., Karfo, P., Nezien, D., Nikiéma, F., Kabre, E., Zongo, C. and Savadogo, A. 2022. Molecular Characterization and the antimicrobial resistance profile of Salmonella spp. isolated from ready-to-eat foods in Ouagadougou, Burkina Faso. Int. J. Microbiol. 2022, 9640828. Wang, D., Zhang, L., Zhou, X., He, Y., Yong, C., Shen, M. and Han, B. 2016. Antimicrobial susceptibility, virulence genes, and randomly amplified polymorphic DNA analysis of Staphylococcus aureus recovered from bovine mastitis in Ningxia, China. J Dairy Sci. 99(12), 9560–9569. | ||
| How to Cite this Article |
| Pubmed Style Mhyson AS, Kadhim MS, Faja OM, Alwan MG, Mohammed BJ. Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Vet. J.. 2024; 14(7): 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 Web Style Mhyson AS, Kadhim MS, Faja OM, Alwan MG, Mohammed BJ. Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. https://www.openveterinaryjournal.com/?mno=198579 [Access: January 24, 2026]. doi:10.5455/OVJ.2024.v14.i7.18 AMA (American Medical Association) Style Mhyson AS, Kadhim MS, Faja OM, Alwan MG, Mohammed BJ. Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Vet. J.. 2024; 14(7): 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 Vancouver/ICMJE Style Mhyson AS, Kadhim MS, Faja OM, Alwan MG, Mohammed BJ. Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Vet. J.. (2024), [cited January 24, 2026]; 14(7): 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 Harvard Style Mhyson, A. S., Kadhim, . M. S., Faja, . O. M., Alwan, . M. G. & Mohammed, . B. J. (2024) Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Vet. J., 14 (7), 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 Turabian Style Mhyson, Afrah Sabeeh, Masar Saab Kadhim, Orooba Meteab Faja, Merriam Ghadhanfar Alwan, and Basima Jasim Mohammed. 2024. Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Veterinary Journal, 14 (7), 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 Chicago Style Mhyson, Afrah Sabeeh, Masar Saab Kadhim, Orooba Meteab Faja, Merriam Ghadhanfar Alwan, and Basima Jasim Mohammed. "Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples." Open Veterinary Journal 14 (2024), 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 MLA (The Modern Language Association) Style Mhyson, Afrah Sabeeh, Masar Saab Kadhim, Orooba Meteab Faja, Merriam Ghadhanfar Alwan, and Basima Jasim Mohammed. "Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples." Open Veterinary Journal 14.7 (2024), 1689-1700. Print. doi:10.5455/OVJ.2024.v14.i7.18 APA (American Psychological Association) Style Mhyson, A. S., Kadhim, . M. S., Faja, . O. M., Alwan, . M. G. & Mohammed, . B. J. (2024) Molecular identification, virulence, and antibiotic resistant genes characteristics of Staphylococcus spp., isolated from milk samples. Open Veterinary Journal, 14 (7), 1689-1700. doi:10.5455/OVJ.2024.v14.i7.18 |

