




| Research Article | ||
Open Vet. J.. 2025; 15(7): 3124-3135 Open Veterinary Journal, (2025), Vol. 15(7): 3124-3135 Research Article Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tractMaha Abdul-Hadi Abdul-Rida Al-Abdula*, Eman Faisal Albghdady, Iman Ibrahim Al Hachem, Abdulrazzaq Baqer and Hazem AlmhannaDepartment of Anatomy and Histology, College of Veterinary Medicine, University of Al-Qadisiyah, Iraq *Corresponding Author: Maha Abdul-Hadi Abdul-Rida Al-Abdula, Department of Anatomy and Histology, College of Veterinary Medicine, University of Al-Qadisiyah, Al-Qadisiyah, Iraq. Email: maha.alabdula [at] qu.edu.iq Submitted: 06/02/2025 Revised: 19/06/2025 Accepted: 27/06/2025 Published: 31/07/2025 © 2025 Open Veterinary Journal
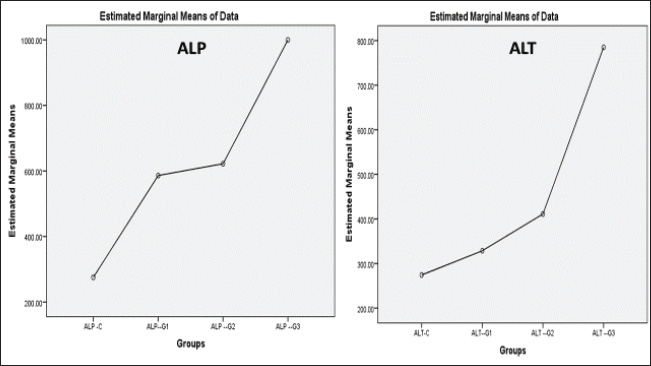
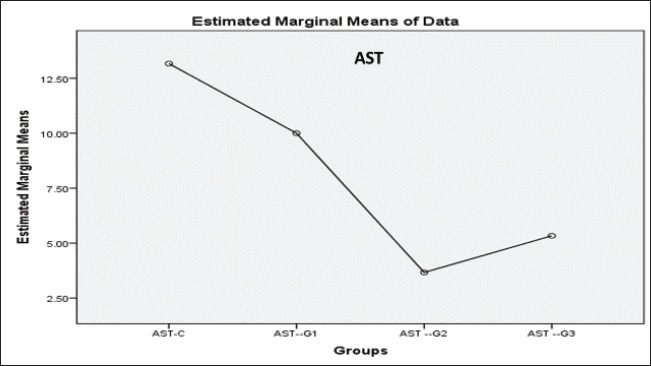
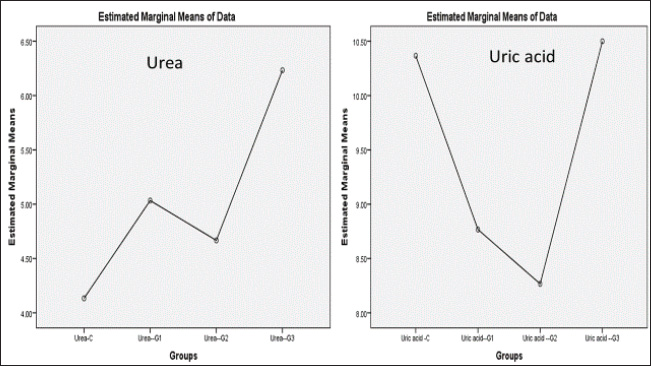
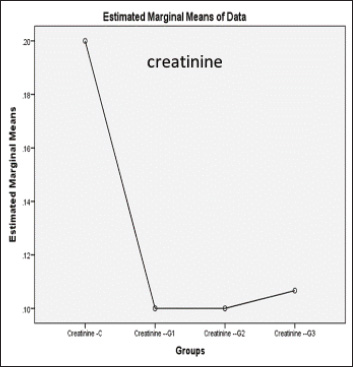
ABSTRACTBackground: Formalin can be very harmful to the health of the quail’s respiratory tract. Aim: This investigation pioneered the examination of formalin gas’s harmful impact on the gene expression of tumor protein TP53 within the quail respiratory system. Methods: In this study, three groups were exposed twice daily 2-hour to formalin gas for 30 days, and the control group did not receive formalin exposure. Tissue samples were collected from the lung, trachea, and blood samples for intervals (10, 20, and 30 days), for gene expression, histological, and biochemical analyses. Results: Our findings showed significant disturbances in liver and kidney function, alongside pronounced pathological alterations in the lungs and trachea, characterized by acute pneumonia. Particularly, 5% formalin gas exposure consistently prompted an increase in the upregulation of TP53 gene expression in both lung and tracheal tissues over the exposure period of formalin. Conclusion: These outcomes strongly suggest that TP53 is a valuable biomarker for assessing formalin gas exposure in both humans and mammals. The results also highlight the significant influence of 5% formalin gas on TP53 gene expression within the respiratory system of quails, with a clear dependence on the length of exposure. Consequently, this study confirmed the potential of evidence that TP53 gene expression is a biomarker for assessing the carcinogenic risks associated with formalin gas exposure in both humans and animals. Keywords: Quail, Formalin, Gene expression TP53, Respiratory system. IntroductionThe quails are a small bird considered an important protein source for humans, such as eggs and meat, because they have a high metabolism efficiency and low cost for breeding compared with industrial poultry and more disease tolerance (Jurafsky, 2014). It is used for conducting many scientific experiments to evaluate the effects of risk diseases, poisons, and drug applications in the poultry industry. These birds easily breed up and tolerate hard conditions compared to other birds (Snow et al., 2005). Formalin (CH2O) is a gas that burns at room temperature with high toxicity and is colorless. Consequently, formalin is considered to have a germicidal effect. It has many uses that make it in demand by consumers, such as its use in manufacturing paper, fertilizers, and paper and fixation of animal tissue for teaching, investigations, and research (Agarwal and Garg, 1983). Much research confirmed that formalin can be used as a food preservative and included in the ingredients of medicines, antiseptics, and cosmetics (Agteren et al., 1998). However, exposure to formalin gas and liquid causes irritation all over the body, especially the eyes and the upper and lower respiratory tract at high or low concentrations, and for long or short periods, especially in birds, for enhancing pigeons (Hasan et al., 2021), ducks (Kuethe, 1988), and quails (Eladl et al., 2022), which leads to suffocation, coughing, and lacrimation. The TP53 gene encodes the tumor suppressor protein TP53, which is a key regulator of cell growth in humans (Kumar et al., 2011). This protein plays a critical role in preventing the development of cancer by controlling the expression of genes involved in cell cycle arrest, DNA repair, and apoptosis (Söderberg-Nauclér, 2006). The dysregulation of p53 expression or function can lead to uncontrolled cell proliferation and subsequent tumor formation (Gupta et al., 2019). Several studies have reported that cellular stress can induce DNA damage, triggering the activation of the tumor suppressor protein TP53 (Zhan, 2005). Moreover, even these conditions, as severe, stress, and damaging DNA, elicit expression of the p53 gene at the cellular level, followed by an increase in cell cycle checkpoints, DNA repair mechanisms, apoptosis, and senescence (Mallette and Ferbeyre, 2007). These reactions are important because they restrict the proliferation of damaged or abnormal cells, thereby modifying the chances of most very types of cancer (Kastan and Bartek, 2004). In addition, numerous studies have stated that the tumor suppressor gene TP53 is essential for inhibiting the development of many sorts of cancers. In addition, deactivating mutations within the TP53 gene are usually the ones that initiate malignant neoplasms in humans, further emphasizing the important role and direct interaction between tumorigenesis of different organs in the body (Hanel and Moll, 2012; Lory et al., 2014). Therefore, because of the consequences of the details on the role of TP53 protein in cancer pathways, this protein has great potential to focus cancer research activity in the plans (Baugh et al., 2017). We decided to investigate the effects of formalin gas on p53 expression after learning about the preceding works that explore the effects of formalin on p53 expression changes. This research focused on the quail’s ability to express and activate the biomarker gene TP53 when subjected to formalin gas as a means to evaluate the potential harm that gasified formalin induces on the quail. Materials and MethodsExperimental designThis study employed sixteen Japanese quails (Coturnix japonica) for experimental purposes. Healthy adult quails were housed in controlled environmental conditions with optimal feeding, lighting, and ventilation in a dedicated facility. The birds were divided into four groups (Each group consists of four quails): control (C) group: quails maintained under standard conditions without formalin exposure. Experimental groups: Group G1 was exposed to 5% formalin gas twice daily for two hours in the morning and two hours in the evening for 10 consecutive days and then sacrificed. Group G2 was exposed to 5% formalin gas twice daily for two hours in the morning and two hours in the evening for 20 consecutive days and then sacrificed. Group G3 was exposed to 5% formalin gas twice daily for two hours in the morning and two hours in the evening for 30 consecutive days and then sacrificed. Sample collection and analysisSerum and tissue samples (trachea and lungs) were collected from each quail at designated time points. Blood and tissue samples were obtained from the trachea and lungs of each experimental group. Serum analysis was conducted to assess liver and kidney function, specifically evaluating the levels of Aspartate aminotransferase (AST), Alanine transaminase (ALT), and Alkaline phosphatase (ALP) using a Beckman Colter AU480 and a Biorad D10 analyzer. The concentrations of urea, creatinine, and uric acid were determined using a UV-kinetic method with commercially available kits (Human Gesellschaft für Biochemie und Diagnostik mbH, Wiesbaden, Germany). Furthermore, for tissue analysis, the tracheal and lung tissue samples were fixed in 10% neutral buffered formalin for histopathological examination. A portion of each tissue was preserved in TRIzol Reagent for subsequent RNA extraction and real-time quantitative PCR (RT-qPCR) analysis. RNA isolation and cDNA synthesisTotal RNA was extracted from lung and tracheal tissues using the TRIzol Reagent method. Briefly, 250 mg of tissue was homogenized in TRIzol Reagent and centrifuged. Chloroform was added to separate phases, and the aqueous phase containing RNA was recovered. Isopropanol was used to precipitate RNA, followed by washing with ethanol. The RNA pellet was air-dried and resuspended in DEPC-treated water. To remove any contaminating DNA, the isolated RNA was treated with DNase I. In the following step, the purified RNA was used to get cDNA with the help of a reverse transcription kit. The procedure was in line with the instructions provided by the manufacturer. The created cDNA, on the other hand, needed to be determined in terms of its concentration and purity, which was achieved using a spectrophotometer. By the end of the procedure, all prepared cDNA samples were diluted to the same concentration for subsequent RT-qPCR analysis. Quantitative gene expression analysis of eosinophil cationic protein (ECP) in QuailQuantitative RT-PCR (qRT-PCR) was used to measure ECP mRNA transcript levels in quail. The following primers were synthesized for the target gene TP53 (XM_015855944.1), which resulted in a 136-bp fragment being amplified (forward: AGCCGCGTTTTAAACTGTGC, reverse: CAAAGTGGCTCTGGAAAGAAGC) and for the housekeeping gene (XM_015873412.2), where a 77-bp fragment was amplified (forward: TGCTGGCATTGCACTGAATG, reverse: CACGGTTGCTGTATCCAAACTC). The qRT–PCR reactions were performed using SYBR green qPCR master mix on a Bio-Rad thermocycler (Hercules, USA). The program settings included an initial step for pre-melting reverse transcription at 50 degrees for 60 minutes, which was followed by 45 cycles of the following: (a) 20 second 95-degree denaturation, (b) 30 second 60 degree annealing/extension, and (c) melting curve analysis from 60 to 95 degrees. For each of the TP53 target genes tested, relative qPCR expression levels were compared with the expression of the control housekeeping gene GAPDH. Histological processingTissue samples of the trachea and lungs were collected and immersed in 10% neutral buffered formalin for 48 hours for fixation. Subsequent histological processing followed standard procedures (Luna, 1968). This process included a graded ethanol series for dehydration, clearing in xylene, and embedding of paraffin. Tissue sections (5–6 μm thick) were obtained using a microtome and stained with hematoxylin and eosin (H&E) for routine histological evaluation. Microscopic examination and image acquisition were performed using an Olympus CH-2 Phase Contrast Microscope (Japan) to analyze the histological features of the trachea and lung tissues in all experimental groups, including the control group. Statistical analysisGene expression levels were quantified using the 2(– Delta Delta C(T)) method (Hellemans et al., 2007). Raw data were subjected to statistical analysis using one-way ANOVA followed by post hoc tests to determine important differences between groups. Statistical significance was set at p < 0.05. All data are presented as mean ± SE. Ethical approvalConducted in accordance with the ethical guidelines for animal research established by the College of Veterinary Medicine, University of Al-Qadisiyah (Approval No. RT324, 2023), following international standards for animal condition and use. ResultsDuring this experiment, the clinical signs of quails under 5% formalin gas were observed, so the birds mostly closed their eyes due to irritation from the formalin gas and displayed nervous signs, depression, anxiety, persistent cough, and lack of appetite, and recorded gradual weight loss. At the same time, the control group showed no clinical signs and grew up healthy. The birds were weighed every 10 days for all groups; thus, the first group’s mean weight was 200 ± 0.66 g, the second group was 188 ± 1.86 g, the third group was 154 ± 0.004 g, and while control was 220 ± 0.013. Significant differences in weight were observed across the four groups. The fourth group showed highly significant differences compared with the rest at p < 0.05. Biochemical study of liver and kidney functionCommon liver enzymes in the quails involving ALP, ALT, and AST, were evaluated after 10 days in each group. Thus, the results showed that ALP and ALT levels were dramatically elevated with exposure days of 5% formalin gas in the experimental groups compared with the control group. In contrast, AST levels were lower in the experimental and control groups. Thus, our results found that levels of ALP in the control and first groups for 10, 20, and 30 days accounted for 275.77 ± 17.21, 586 ± 55.7, 622 ± 69.5, and 999.6 ± 87.6, respectively, (Fig. 1), while the control and second groups for ALT were accounted for 274.44 ± 8.23, 328 ± 60.3, 411 ± 84.4, and 785 ± 265, respectively, (Fig. 2). In contrast, the control and third groups for AST accounted for 13.16 ± 0.7, 10 ± 5.7, 5.33 ± 0.7, and 3.33 ± 1.5, respectively, (Fig. 2). Consequently, there were statistically significant differences for both ALP and ALT compared to the rest of the groups (p < 0.05), in contrast to AST, which showed a significant decline compared to the control group and the other groups (p < 0.05). The levels of urea, uric acid, and creatinine in the blood of the quails were also assessed during this experiment after every 10 days for the control and groups. Thus, the results showed that the levels of urea, uric acid, and creatinine ascending with days of exposure to 5% gas formalin. This indicated that formalin gas affected kidney function. As a result, the control and first groups for urea at 10, 20, and 30 days accounted for 4.13 + 0.23, 4.66 + 0.3, 5.08 + 0.80, and 6.23 + 0.68, respectively, (Fig. 3), while the control and second groups for uric acid were accounted for10.36 + 1.50, 8.76 + 0.70, 8.26 + 0.6, and 8.20 + 0.4, respectively (Fig. 2). In contrast, the control and third groups for creatinine accounted for 0.2 + 0.1, 0.1 + 0.0, 0.1 + 0.00, and 0.1 + 0.00, respectively (Fig. 4). There were statistically significant differences for both urea and uric acid compared with the rest of the groups (p < 0.05), in contrast to creatinine, which showed a significant decline compared with the control group and the other groups. Histological and histopathological studyThe histological study focused on the trachea and lung tissue of the quail during the experiment. Accordingly, sections of the normal tissue of the trachea in the control group showed that it was lined by a thin mucosal layer and supported by overlapping-shaped rings of cartilage and the smooth and skeletal muscle outside the trachea. The epithelial layer was a pseudostratified ciliated columnar epithelium with numerous mucous glands and the goblet cells of mucosal epithelia. The trachea is lined with pseudostratified ciliated columnar epithelium containing numerous mucous glands. In the formalin gas group, 5% of the trachea showed tracheal loss of cilia from the epithelia and more spaces and gaps between the mucosa, submucosal layer, and cartilaginous layer (Figs 5, 6A). Similarly, the lung tissue of the control quail exhibited branching of the left and right bronchi and parabronchi, encircled by numerous blood vessels. The parabranchial tubes opened into the atria and were surrounded by smooth muscle. The atrial goon appeared as an infundibula. These infundibular ducts are formed by a complex air capillary with oval shapes. Moreover, a simple squamous epithelium lined the parabronchi, atria, infundibula, and air capillaries. In the three groups of birds exposed to 5% formalin gas, the lung tissue of the quail displayed acute inflammation, evidence of interstitial pneumonia, and severe pulmonary edema, in addition to the alveoli were filled with secretions of the inflammatory, and infiltration of immune cells. The bronchial area was congested with secretions, and the bronchial wall was thickened with an accumulation of inflammatory secretions and red blood balls (Figs 5, 6B).
Fig. 1. Histogram of liver enzymes shows: that ALP and ALT values in the third groups were significant increase at P<0.05, and more than first and second groups as well as control groups
Fig. 2. Histogram of liver enzymes shows that the AST value in the third group was significantly decreased at P<0.05, less than in the first and second groups and the control groups.
Fig. 3. Histogram of urea and uric acid levels show: that urea and uric acid values in the blood of the third groups were highly significant at P≥ 0.05 more than those of the first and second groups as well as the control groups.
Fig. 4. Histogram of creatinine value in blood shows: creatinine value in the third groups was highly significant at P<0.05 decreased more than in the first and second groups as well as in the control groups.
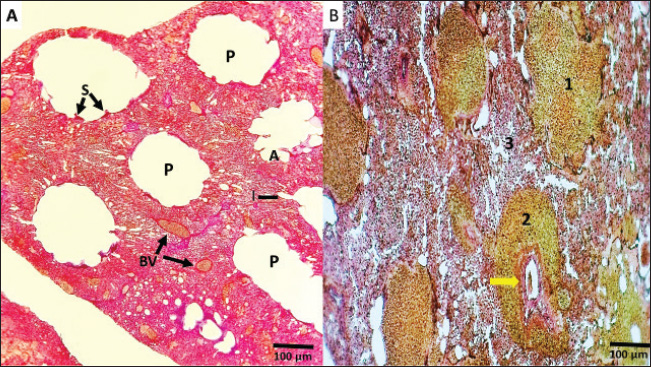
Fig. 5. A and B. Histological sections of the lung show: (A) in the control lung, the parabronchi (P) are encircled by numerous blood vessels (BV); the parabranchial tubes open into the atria(A) surrounded by smooth muscle (S). The atrial goon as ducts called infundibula (I) B: Section of the lung showed severe chronic interstitial pneumonia, severe pulmonary edema was evident, the alveoli were filled with exudate and inflammatory cells (1), engorgement of bronchiole with exudate (yellow arrow), and thickening of the bronchiolar wall with accumulation of inflammatory cells (2), severe interstitial thickening and infiltration of inflammatory cells (3) H&E, (A, B) (Magnificence, 200X).
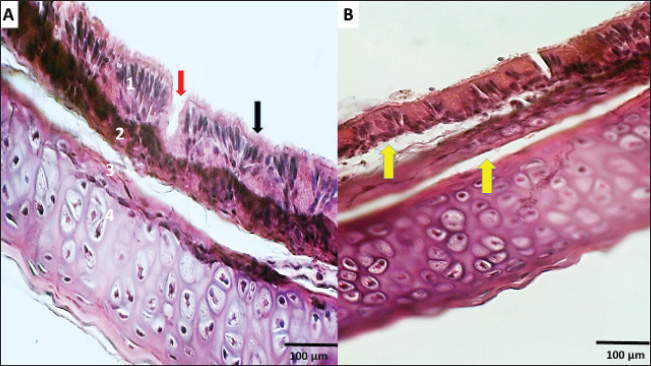
Fig. 6. (A) Section of normal trachea showing the mucosa (1) containing goblet cells (red arrow) and pseudostratified columnar ciliated epithelia tissue (black arrow), lamina propria and mucosa muscularis (2), and hyaline cartilage (3). (B) section shows detachment between the mucosa, submucosa, and cartilaginous layer (yellow arrows). H&E (A&B), (Magnificence, 400X. Real-time quantitative PCR (RT-qPCR) findingsOur investigation utilizing RT-qPCR demonstrated variations in the amplification curves of the TP53 gene within tracheal and lung tissues across three experimental groups: those exposed to 5% formalin gas for different durations (10, 20, and 30 days) and a control group (Figs. 7, 9, and 11). The threshold cycle (Ct) values, which are indicative of gene expression levels, exhibited significant differences between the groups. Melting curve analysis of the RT-qPCR primers confirmed the high specificity of TP53 gene detection, with no evidence of nonspecific amplification products (Figs. 8, 10, and 12). The melting peak temperatures for TP53 ranged from 70 °C to 80 °C. These findings suggest that exposure to 5% formalin gas can upregulate TP53 expression in both tracheal and lung tissues, with increasing expression levels observed over the exposure period (p < 0.05). In contrast, in the trachea, TP53 expression levels were significantly elevated after 10, 20, and 30 days of formalin gas exposure compared with the control group. However, no significant differences in TP53 expression were observed between the tracheal tissue exposure groups. Conversely, in lung tissue, TP53 expression in the group exposed to formalin gas for 30 days (third group) was significantly higher than in the group exposed for 10 days (first group) (p=0.042) and the control group (p=0.018). Although the 30-day group showed a trend toward higher TP53 expression compared with the 20-day group (second group), this difference did not reach statistical significance (p=0.072). DiscussionMany studies have shown that low formalin concentrations in closed, poorly ventilated environments can reduce blood oxygen saturation levels. Hypoxia stimulates the stimulation of metabolic activity in the body and increases energy requirements (Hun et al., 2010). Formalin can arguably be used for longer periods of time as the body generates significant vasodilation in return for the lack of oxygen in the tissues (Liu et al., 2018). Therefore, our study was conducted to observe the impact of 5% formalin gas on the expression of TP53 within the respiratory system of quail. Our findings revealed that quails exposed to 5% formalin gas displayed obvious clinical signs, including anxiety, depression, and elevated stress levels, during the period of exposure time. These observants reported by (Follett and Maung, 1978; Awbi, 2003; Salthammer et al., 2010), who found that formalin gas exposure could prompt tissue hypoxia and lead to specific clinical signs during the exposure period. Importantly, our study distinguished significant changes in liver enzyme concentrations. Thus, ALP and ALT concentrations were clearly observed upregulation increase with exposure time, whereas AST levels declined compared with the control group. These enzyme alterations provide potentially evidence that formalin exposure might be caused by hepatic damage, depending on the severity of liver tissue damage. An earlier study has confirmed that hepatocellular injury is widely associated with increased levels of ALP, ALT, and AST liver enzymes (Khan et al., 2006), which supports our observations. Moreover, these enzyme changes for liver cell function are evident and agree with our outcomes, which detected increasing levels of ALP and ALT in groups exposed to formalin gas compared with the control group. More specific studies have investigated the effect of exposure to liquid formalin in drinking water at higher concentrations on quails and noticed negative significance impression liver function (Thiele et al., 1999; Vahdatpou et al., 2011) and decreased activity of key liver enzymes, including ALP, AST, and ALT. However, AST was dramatically decreased in groups exposed to formalin gas in comparison compared to the control group. This might be because different routes of formalin exposure led to alterations in liver cell function. Significantly, our findings confirmed these previous observations, suggesting that both liquid and gaseous formalin exposure can significantly alter liver enzyme activity in birds. This study further demonstrated that different routes of formalin exposure can elicit distinct effects on liver function, as evidenced by significant differences in ALP, AST, and ALT levels between the experimental and control groups. Furthermore, prior researchers have shown that glomerulus damage in the quail can induce a decrease in filtration rates and affect urea reabsorption through the tubule (Echols, 2006). Therefore, dehydration, fever, and markedly increased tubular sodium reabsorption may cause an increase in the urea/creatinine ratio and uric acid (Kasmani et al., 2012). Our observations confirm the previous data. We noticed pronounced kidney malfunctions caused by exposure of quail to 5% formalin gas over a duration of increasing gas concentrations. These changes were associated with significant increments in blood urea, uric acid, and creatinine levels in the experimental groups. Thus, these results suggest that formalin gas exposure under all conditions worsens kidney malfunction in the quail. Formaldehyde is a highly toxic compound with the potential to insult different body organs, particularly the respiratory system, including the trachea and lungs (Golden and Valentini, 2014). Our study revealed severe pathological alterations in the trachea and lung tissue of quails exposed to formaldehyde for a long period. However, as in any other studies concerning humans or murine models (Pope et al., 2002; Risom et al., 2005; Eladi et al., 2022), long-term exposure to formaldehyde gas increases the risk of lung cancer. This research aimed to elucidate the up-regulation of the TP53 gene in the tracheal and lung tissues of quail under the egis of a 5% formaldehyde gas exposure over different time periods.
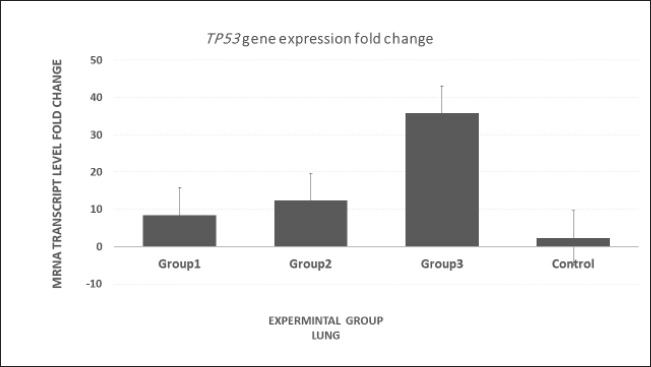
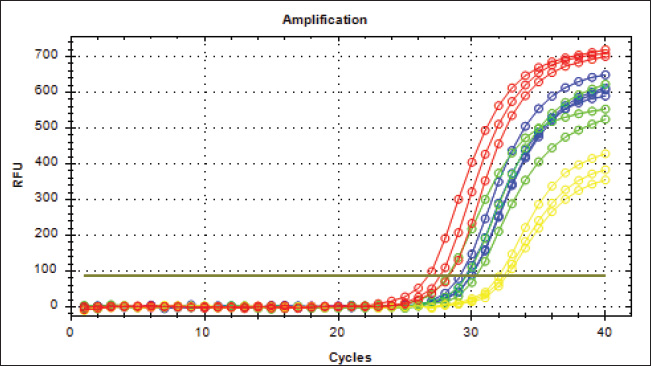
Fig. 7. mRNA expression analysis of TP53 in lung tissue revealed significantly elevated levels in Group 3 compared with Groups 1, 2, and the control group.
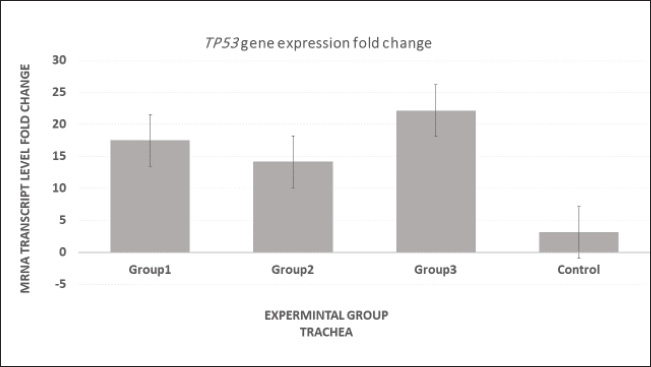
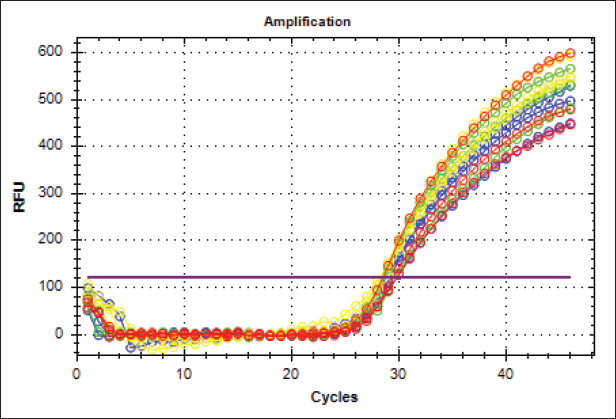
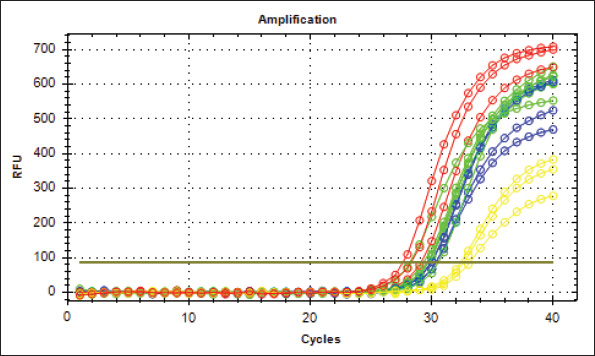
Fig. 8. Analysis of TP53 gene expression in the trachea revealed that mRNA p53 levels were significantly elevated in the third group compared with the other two groups and the control group.
Fig. 9. Real-Time PCR Amplification Plots of TP53 Gene Expression in Lung Tissue. The green plots (G1 group), the Blue plots (G2 group), the red plots (G3 group), and the yellow plots (Control group).
Fig. 10. Real-Time PCR amplification plots of the housekeeping gene Gapdh in lung tissue samples from four experimental groups: G1 (green), G2 (blue), G3 (red), and Control (yellow).
Fig. 11. TP53 gene amplification plots (Real-Time PCR) from tracheal tissue samples. G1: Green, G2: Blue, G3: Red, Control: Yellow.
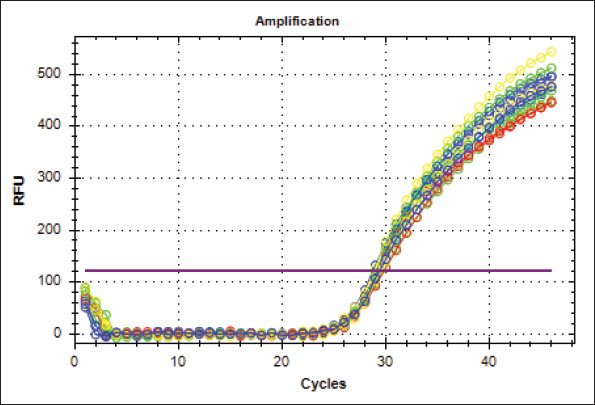
Fig. 12. This figure presents the amplification curves obtained using real-time PCR of the housekeeping gene Gapdh in tracheal tissue samples. The green curves are from the G1 group, the blue curves are from the G2 group, the red curves are from the G3 group, and the yellow curves are from the control groups. The P53 pathway has been shown to play essential roles in regulating apoptosis in tracheal lung cells, thereby further influencing processes in the Url 7 Thep53-dependent apoptotic pathway (Surien et al., 2021). Thus, our results were able to reveal that the P53 protein expression in the trachea and lungs increased significantly following quail exposure to formaldehyde, suggesting that formaldehyde-induced apoptosis may involve both P53-dependent and non-P53-dependent mechanisms. Furthermore, these findings emphasize the importance of considering the temporal aspects of formaldehyde exposure, including the frequency and duration of exposure periods, to understand its physiological effects on the respiratory system. ConclusionThis investigation revealed that exposure to 5% formalin gas resulted in substantial disruption of liver and kidney function. In particular, prolonged exposure upregulated TP53 gene expression in the trachea and lungs. This persistent elevation of TP53 gene expression may indicate abnormal tissue responses, leading to a high probability of mutagenicity or carcinogenesis. Thus, TP53 should be examined as a significant biomarker for effectively evaluating exposure to formalin gas in the respiratory systems of animals and humans. AcknowledgmentsThe authors thank the College of Veterinary Medicine, University of Al-Qadisiyah, Iraq. FundingNo specific funding was received. Authors’ contributionsAll authors have participated equally in the current research. Conflict of interestThe conflict of interest is not declared. Data availabilityThe data are available via the corresponding author when requested. ReferencesBanerjee, S., Tsutsui, K. and Chaturvedi, C.M. 2016. Apoptosis-mediated testicular alteration in Japanese quail (Coturnix coturnix japonica) in response to temporal phase relation of serotonergic and dopaminergic oscillations. J. Exp. Biol. 219, 1476–1487. Baugh, E.H., Ke, H., Levine, A.J., Bonneau, R.A. and Chan, C.S. 2018. Why are there hotspot mutations in the TP53 gene in human cancers. J. Cell Death Differ. 25, 154–160. Broom, D.M. and Corke, M.J. 2002. Effects of disease on farm animal welfare. Acta Vet. Brno. 71, 133–136. Echols, M.S. 2006. Evaluating and treating the kidneys. In: Harrison and lightfoot clinical avian medicine. New York City: Pix Publishing, Inc., pp: 451–492. Eladl, A.H., Farag, V.M., El-Shafei, R.A., Aziza, A.E., Awadin, W.F. and Arafat, N. 2022. Immunological, biochemical, and pathological effects of vitamin C and Arabic gum co-administration on H9N2 avian influenza virus vaccinated and challenged laying Japanese quails. BMC Vet. Res. 18, 1–14. Follett, B.K. and Maung, S.L. 1978. With gonadotrophin and testosterone levels, the testicular maturation rate in quail was exposed to various artificial photoperiods and natural day lengths. J. Endocrinol. 78, 267–280. Golden, R. and Valentini, M. 2014. Formaldehyde and methylene glycol equivalence a critical assessment of chemical and toxicological aspects. Regul. Toxicol. Pharmacol. 69, 178–186. Grigorev, I.P. and Korzhevskii, D.E. 2018. Current technologies for fixation of biological material for immunohistochemical analysis (in Russian). J. Mid Technol. 10, 156–164. Grillo, F., Pigozzi, S., Ceriolo, P., Calamaro, P., Fiocca, R. and Mastracci, L. 2015. Factors affecting immunoreactivity in long-term storage of formalin-fixed paraffin-embedded tissue sections. Histochem. Cell Biol. 144, 93–99. Gupta, A., Shah, K., Oza, M.J. and Behl, T. 2019. Reactivation of TP53 gene by MDM2 inhibitors a novel therapy for cancer treatment. Biomed. Pharmacother. 109, 484–492. Hanel, W. and Moll, U.M. 2012. Links between mutant TP53 and genomic instability. J. Cell Biochem. 113, 433–439. Hasan, I., Pervin, M., Kobir, M.A., Sagor, S.H. and Karim, M.R. 2021. Effect of formaldehyde and urea contaminated feed exposure into the liver of young and adult pigeons (Columba livia). Vet. World 14, 769–787. Hellemans, J., Mortier, G., De Paepe, A., Speleman, F. and Vandesompele, J. 2007. Base relative quantification framework and software for management and automated real-time quantitative PCR data analysis. Genome Biol. 8, 1–14. Herrnstein, R.J., Loewenstein, G.F., Prelec, D. and Vaughan, W. 1993. Utility maximization and melioration: Internalities in individual choice. J. Behav. Decis. Mak. 6, 149–185. Hun, D.E., Corsi, R., Morandi, T. and Siegel, J.A. 2010. Formaldehyde in residences: long-term indoor concentrations and influencing factors. Indoor. Air 20, 196–203. Kasmani, F.B., Torshizi, M.K., Allameh, A. and Shariatmadari, F. 2012. A novel aflatoxin-binding Bacillus probiotic: performance, serum biochemistry, and immunological parameters in Japanese quail. Poult. Sci. 91, 1846–1853. Kastan, M.B. and Bartek, J. 2004. Cell-cycle checkpoints and cancer. Nature 432, 316–323. Khan, A., Hussain, S.M. and Khan, M.Z. 2006. Effects of formalin feeding or administering into the crops of white leghorn cockerels on hematological and biochemical parameters. Poult. Sci. 85, 1513–1519. Kuethe, D.O. 1988. Fluid mechanical valving of airflow in bird lungs. J. Exp. Biol. 136, 1–12. Kumar, M., Lu, Z., Takwi, A., Chen, W., Callander, N.S., Ramos, K.S. and Li, Y. 2011. Negative regulation of the tumor suppressor p53 gene by microRNAs. Oncogene 30, 843–853. Leroy, B., Girard, L., Hollestelle, A., Minna, J.D., Gazdar, A.F. and Soussi, T. 2014. Analysis of TP53 mutation status in human cancer cell lines A reassessment. Hum. Mutat. 35, 756–765. Liu, J., Dai, X., Li, X., Jia, S., Pei, J., Sun, Y. and Jian, Y. 2018. Indoor air quality and occupants’ ventilation habits in China seasonal measurement and long-term monitoring. Build Environ. 142, 119–129. Livak, K.J. and Schmittgen, T.D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta). Res. Methodol. 25, 402–410. Mallette, F.A. and Ferbeyre, G. 2007. The DNA damage signaling pathway connects oncogenic stress to cellular senescence. Cell Cycle 6, 1831–1836. Pope, C.A., Burnett, M., Thun, E.E., Calle, D., Krewski, K. and Ito, G.D. 2002. Lung cancer, cardiopulmonary mortality, and long-term fine particulate air pollution exposure. JAMA 287, 1132–1141. Risom, L., Møller, P. and Loft, S. 2005. Oxidative stress-induced DNA damage by particulate air pollution. Mutat. Res. Fund Mol. Mech. Mutagen 592, 119–137. Salthammer, T., Mentese, S. and Marutzky, R. 2010. Formaldehyde in the indoor environment. Chem. Rev. 110, 2536–2572. Snow, A.A., Andow, D.A., Gepts, P., Hallerman, E.M., Power, A., Tiedje, J.M. and Wolfenbarger, L. 2005. Genetically engineered organisms and the environment: current status and recommendations. Ecol. Appl. 15, 377–404. Söderberg-Nauclér, C. 2006. Does cytomegalovirus play a causative role in developing various inflammatory diseases and cancer. J. Intern. Med. 259, 219–246. Surien, O., Ghazali, A.R. and Masre, S.F. 2021. Chemopreventive effects of pterostilbene through p53 and cell cycle in mouse lung of squamous cell carcinoma model. Sci. Rep. 11, 1–11. Thiele, G.M., Miller, J.A., Klassen, L.W. and Tuma, D.J. 1999. Chronic ethanol consumption impairs receptor-mediated endocytosis of formaldehyde-treated albumin by isolated rat liver endothelial cells. Hepatology 29, 1511–1517. Vahdatpour, T., Nikpiran, H., Babazadeh, D., Vahdatpour, S. and Jafargholipour, M.A. 2011. Effects of Protexin®, Fermacto®, and their combination on blood enzymes and performance of Japanese quails (Coturnix japonica). Ann. Biol. Res. 2, 283–291. Zhan, Q. 2005. Gadd45a, TP53- and BRCA1-regulated stress protein, in the cellular response to DNA damage. Mutat. Res. Fund Mol. Mech. Mutagen 569, 133–143. | ||
| How to Cite this Article |
| Pubmed Style Al-abdula MAA, Albghdady EF, Hachem IIA, Baqer A, Almhanna H. Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Vet. J.. 2025; 15(7): 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 Web Style Al-abdula MAA, Albghdady EF, Hachem IIA, Baqer A, Almhanna H. Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. https://www.openveterinaryjournal.com/?mno=241524 [Access: January 25, 2026]. doi:10.5455/OVJ.2025.v15.i7.24 AMA (American Medical Association) Style Al-abdula MAA, Albghdady EF, Hachem IIA, Baqer A, Almhanna H. Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Vet. J.. 2025; 15(7): 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 Vancouver/ICMJE Style Al-abdula MAA, Albghdady EF, Hachem IIA, Baqer A, Almhanna H. Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Vet. J.. (2025), [cited January 25, 2026]; 15(7): 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 Harvard Style Al-abdula, M. A. A., Albghdady, . E. F., Hachem, . I. I. A., Baqer, . A. & Almhanna, . H. (2025) Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Vet. J., 15 (7), 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 Turabian Style Al-abdula, Maha Abdul-hadi Abdul-rida, Eman Faisal Albghdady, Iman Ibrahim Al Hachem, Abdulrazzaq Baqer, and Hazem Almhanna. 2025. Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Veterinary Journal, 15 (7), 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 Chicago Style Al-abdula, Maha Abdul-hadi Abdul-rida, Eman Faisal Albghdady, Iman Ibrahim Al Hachem, Abdulrazzaq Baqer, and Hazem Almhanna. "Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract." Open Veterinary Journal 15 (2025), 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 MLA (The Modern Language Association) Style Al-abdula, Maha Abdul-hadi Abdul-rida, Eman Faisal Albghdady, Iman Ibrahim Al Hachem, Abdulrazzaq Baqer, and Hazem Almhanna. "Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract." Open Veterinary Journal 15.7 (2025), 3124-3135. Print. doi:10.5455/OVJ.2025.v15.i7.24 APA (American Psychological Association) Style Al-abdula, M. A. A., Albghdady, . E. F., Hachem, . I. I. A., Baqer, . A. & Almhanna, . H. (2025) Investigation of formalin gas-induced alterations in TP53 gene expression in quail respiratory tract. Open Veterinary Journal, 15 (7), 3124-3135. doi:10.5455/OVJ.2025.v15.i7.24 |

