




| Research Article | ||
Open Vet. J.. 2025; 15(9): 4310-4321
Open Veterinary Journal, (2025), Vol. 15(9): 4310-4321 Research Articlewww.eldaghayes.com Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female ratsNurasmi Nurasmi1,2, Wasmen Manalu1*, I Ketut Mudite Adnyane3 and Andriyanto Andriyanto41Division of Physiology, School of Veterinary Medicine and Biomedical Sciences, IPB University, Bogor, Indonesia 2Fisheries Product Technology Department, Faculty of Fishery and Marine Science, University of Borneo, Tarakan, Indonesia 3Division of Anatomy, Histology and Embryology, School of Veterinary Medicine and Biomedical Sciences, IPB University, Bogor, Indonesia 4Division of Pharmacology and Toxicology, School of Veterinary Medicine and Biomedical Sciences, IPB University, Bogor, Indonesia *Corresponding Author: Wasmen Manalu. Division of Physiology, School of Veterinary Medicine and Biomedical Sciences, IPB University, Bogor, Indonesia. Email: wasmenma [at] apps.ipb.ac.id Submitted: 26/04/2025 Revised: 05/08/2025 Accepted: 22/08/2025 Published: 30/09/2025 © 2025 Open Veterinary Journal
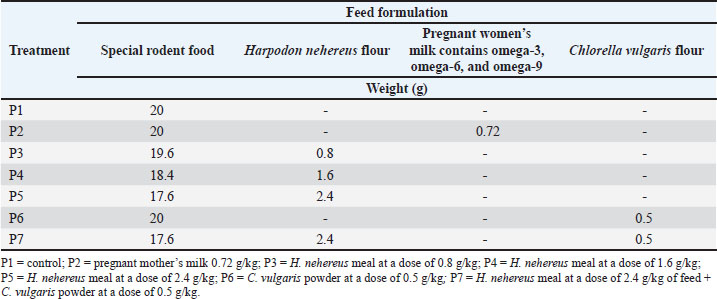
ABSTRACTBackground: Productivity is primarily determined by the ability of the mother to produce healthy, strong, and optimally developed offspring from the zygote to birth, ultimately improving postnatal growth and health. The mother’s health is a primary support system in reproductive health, as reflected in the values of body weight, estrogen and progesterone, uterus, ovaries, and vascularization, all of which are crucial in enhancing reproductive performance. Harpodon nehereus and Chlorella vulgaris have compounds that can enhance and repair reproductive health and performance. Aim: This study aimed to evaluate the effects of H. nehereus and C. vulgaris supplementation on increasing the body weight and performance of female rats from pre-pregnancy to pregnancy. Methods: Nulliparous and sexually mature female Sprague–Dawley rats were subjected to an experimental study with a completely randomized design. Female virgin rats weighing 150–200 g were divided into seven dose groups: P1, P2, P3, P4, P5, P6, and P7. Each group of virgin female rats was given a predetermined treatment dose. The main ingredients of the treatment feed are fishmeal, H. nehereus, and C. vulgaris. The observed variables included body weight, estrogen and progesterone levels, uterus, ovaries, and vascularization. Results: The results showed increased estrogen and progesterone levels in the H. nehereus and C. vulgaris flour treatment group. Supplementation of experimental rats with H. nehereus and C. vulgaris fishmeal increased estrogen and progesterone levels, primary and secondary follicles, and uterine gland development. Conclusion: Additionally, maternal vascularization increased after supplementation treatment. Rats’ body weight before and after pregnancy did not differ significantly across all treatments (p > 0.05). Estrogen and progesterone levels increased in the test animals. The number of primary follicles, secondary follicles, and vascularization significantly differed (p < 0.05) in all treatment groups compared with the control. Keywords: Chlorella vulgaris, Harpodon nehereus, Hormone, Reproductive. IntroductionReproductive health is a global issue that receives international attention. Efforts to improve the health status of mothers and fetuses aim to reduce mortality and the incidence of illness in both mothers and fetuses. The quality of the fetus is determined by the quality of the mother’s process (Manalu, 1999). Currently, stunting is one of the most significant global health issues. Malnutrition can cause stunting, impact intelligence performance, and increase the risk of developing degenerative diseases in adulthood (Jiang et al., 2023; Siddiqa et al., 2024). Malnutrition occurs when the body has an imbalance of nutrients, specifically when the nutrients required by the body are not in balance with the amount available for use (World Health Organization, 2021). Improving maternal reproductive health and developing body organs during prenatal and postnatal growth is necessary for stunting prevention. Adequate nutrition during both prenatal and postnatal growth will enhance maternal health and fetal development. Pregnant women’s nutrition requires attention because it significantly impacts the first 1,000 days of a child’s life (Likhar and Patil, 2023). Efforts to achieve optimum reproductive health and growth can be conducted by improving the health condition of the mother during the period (Andriyanto et al., 2022). One of these improvements is increasing and improving the mother’s reproductive hormone synthesis and secretion. Estrogen and progesterone are key hormones that play essential roles in the reproductive system (Okada et al., 2018). The reproductive hormone estrogen produced by the developing follicles during estrus and progesterone after ovulation and during pregnancy are the primary signals that initiate the reproductive process (Manalu et al., 2000). These pregnancy hormones provide the initial signal to start the comprehensive reproductive process, starting from the preparation of the ovum and sperm, fertilization, preparation of the uterus for implantation, and development of the placenta, embryo, and fetus, which ultimately affects the flow of nutrients to the developing embryo and fetus and gene expression, which will ultimately have implications for birth weight, pre-weaning growth to adulthood. One alternative solution to overcome these problems is to improve the mother’s reproductive system, starting with the improvement of hormones during the pregnancy process using natural ingredients from Harpodon nehereus fish meal and Chlorella vulgaris powder, which contain proteins, steroids, omega-3, omega-6, and omega-9 essential during pregnancy. Nomei (H. nehereus) fish is a soft fish with high economic value that can be used sustainably. Based on data from the Sectoral Statistical Data Information System of North Kalimantan Province, Tarakan City, the Nomei fish has considerable potential, namely 100 tons per month in its fresh form, equivalent to approximately 3 tons of dried Nomei fish. The protein test value of the dried Nomei fish, which results from fishery processing, was 66.52% (Salim et al., 2022). Nomei fish can be a source of protein, omega-3, omega-6, and omega-9. Research on H. nehereus specifically addresses its benefits for improving reproductive health and intelligence; however, no research has been conducted to date. While there may not be any research specifically addressing H. nehereus, there is a wealth of relevant research on marine fish and omega-3 fatty acids. Research has shown that omega-3 fatty acids, including docosahexaenoic acid (DHA), can improve egg quality, which is important for fertility, in women. A study by Gaskins et al. (2016) showed that omega-3 consumption from seafood is associated with improved egg quality and pregnancy outcomes in women and suggests that DHA supplementation during pregnancy may improve fetal brain development and prevent preterm birth. Omega-3s also play a role in reducing the risk of pregnancy complications such as preeclampsia. However, C. vulgaris remains in high demand within the community because it lacks product development and other processing. Chlorella vulgaris is rich in omega-3, omega-6, and omega-9 fatty acids; omega-6 represents more than 60% of the total fatty acids (Paliwal and Jutur, 2021). Chlorella vulgaris contains phenolic compounds, tannins, flavonoids, cardiac glycosides, saponins, and terpenoids. The concentration of omega-3 and omega-6 in C. vulgaris is 55.98%; DHA and eicosapentaenoic acid are 331.07 mg/g DM. Chlorella vulgaris is also rich in beneficial sterols and steroids that are beneficial for the health of reproductive organs and fertility and are utilized as a source of functional food, nutraceuticals, and food supplements (Sathasivam et al., 2019; Wang et al., 2024). The role of algae in the reproductive system is based on its sterol content and the steroid production. Sterols, such as cholesterol, in algae undergo a biochemical process to form steroid derivative compounds during steroid formation, known as steroidogenesis. These two natural ingredients provide nutrients that support hormonal balance, improve cell quality, and minimize oxidative stress that can compromise reproductive health. Chlorella vulgaris provides antioxidants and omega-3 fatty acids, whereas H. nehereus provides protein, DHA, and omega-3s, which help regulate hormones and improve cell health. Both play a vital role in supporting brain development. Chlorella vulgaris provides DHA and eicosapentaenoic acid, which are vital for the nervous system and cognitive development of a fetus. DHA is crucial for early brain development. This study aimed to evaluate the reproductive performance of Sprague-Dawley rats given H. nehereus meal and C. vulgaris powder-based supplements by measuring estrogen and progesterone body weight, hormone levels, ovarian and uterine histology, and vascularization. Materials and MethodsPreparation of Harpodon nehereus and Chlorella vulgaris extractsNomei fish or H. nehereus powder was collected from the Juata Laut, Tarakan, North Kalimantan, Indonesia. The collected Nomei fish were immediately cleaned and washed. The fish were cut into thinner pieces, dried in the sunlight, and then baked in a 60°C oven for 24 hours. The dried fish were then ground to produce fishmeal. Chlorella vulgaris was obtained from SEDUH. The product comes with a certificate that guarantees its safety and quality. Experimental designA total of 42 female, 8-week-old Sprague-Dawley rats (iRATco Veterinary Laboratory Services, Bogor, Indonesia), weighing approximately 150–200 g, in the diestrus phase of the estrus cycle, were divided equally into seven different treatment groups based on the doses of treatment. This study was designed to evaluate the effects of H. nehereus and C. vulgaris flour mixed with commercial feed at various doses on experimental rats. Commercial rat feed was used for the mixture. The supplementation study was conducted 20 and 23 days before conception and 23 days during gestation. The supplementation study on the experimental animals before termination was conducted for 43 days. The doses were divided into seven groups (Table 1): (i) experimental rats fed with Ratbio (PT). Japfa Comfeed. Sidoarjo, East Java, Indonesia) without supplementation of H. nehereus and C. vulgaris (P1 group as a control group); (ii) The experimental rats fed ration supplemented with pregnant milk (milk for pregnant mother) at a dose of 0.72 g/kg feed (Prenagen, PT Kalbe Farma Tbk) without supplementation of H. nehereus and C. vulgaris (P2 group); (iii) Experimental rats fed ration supplemented with H. nehereus at a dose of 0.8 g/kg feed without C. vulgaris supplementation (P3 group); (iv) Experimental rats fed a ration supplemented with H. nehereus at a 1.6 g/kg feed without C. vulgaris supplementation (P4 group); (v) Experimental rats fed a ration supplemented with H. nehereus at a 2.4 g/kg feed without C. vulgaris supplementation (P5 group); (vi) Experimental rats fed a ration supplemented with C. vulgaris at 0.5 g without H. nehereus supplementation (P6 group); and (vii) Experimental rats fed a ration supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7 group). Table 1. Feed formulation.
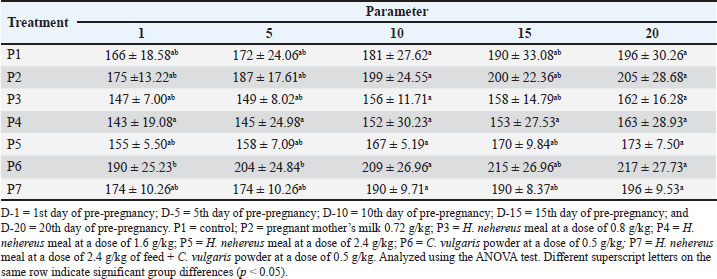
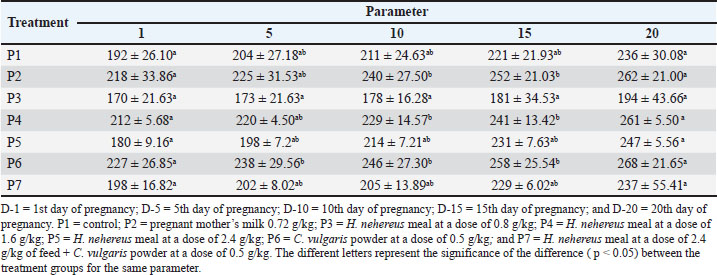
The feed formulation was supplemented at different doses (Table 1). The experimental rats were fed 10% of their body weight. The rats were supplemented for 20 days before mating and until delivery. On the first day of delivery, the mother rats were sacrificed for uterus and ovary collection. After observing each experimental rat’s estrus cycle, pregnancy was induced by confining the female rat overnight with a fertile male rat. The presence of spermatozoa in the vaginal smear was defined as pregnancy (D 0) on the morning of the next day. Then, 24 rats became pregnant in each group according to the assigned treatment doses. All experimental rats were euthanized using ketamine (100 mg/kg of body weight [BW]) (Ket-A-100®, Agrovet Market, Canada) and xylazine (3 mg/kg of BW) (Xyla, Interchemie, Netherlands), followed by perfusion as described previously (Ko et al., 2019). The experimental rats were maintained according to the standard guidelines for animal care and use (National Research Council, 2011). They were provided an ad libitum standard rodent diet and access to drinking water. The number of rat pups born was recorded. Pre-pregnancy and pregnancy weight gain measurement of the motherPre-pregnancy weight gain was calculated by subtracting the final weight from the initial weight of the treated rat. Weight gain was measured every 5 days before and during pregnancy. The weight gain during pregnancy was calculated by subtracting the initial weight from the final weight. Weighing was performed using a mini digital scale (MH-Series Pocket Scale) with a precision of 0.01g to obtain birth weight data. Analysis of estrogen and progesterone levelsBlood samples of the maternal experimental rats were collected through the lateral tail vein as much as 3 ml one day before mating (D-0), day 5 of pregnancy (D-5), day 10 of pregnancy (D-10), and day 15 of pregnancy (D-15). Serum was separated by centrifugation (3,500 × g, 10 minutes) and stored at −20°C until further analysis. The concentrations of estrogen in the serum of maternal rats were determined using a Rat Estrogen ELISA Kit (FineTest, Cat no. ER1505, China), and serum progesterone concentrations were determined using a Pg ELISA Kit (FineTest, Cat no. ER1255, China) according to the manufacturer’s instructions. The optical density was measured using an ELISA reader (TC 96 microplate reader, Teco Company, Canada). Uterine analysisBefore euthanasia, a picture of the uterus was taken with an Olympus stereo microscope SD30 (Olympus, Japan) and a Canon EOS D1100 SLR camera (Canon, Taiwan) for vascular analysis. We analyzed the length of the left and right uterine cornua, the diameter of the uterine cornua, and the proportion of the uterine blood vessel area to the total uterine area using ImageJ software (National Institutes of Health, US). The proportion of uterine blood vessels was calculated using the following formula: Histology of the ovaries and uterusThe ovaries and uteruses were collected from newborns and fixed in 10% buffered formalin, dehydrated, and embedded in paraffin. Paraffinized tissue blocks were sectioned into 4-μm-thick sections using a Leica RM2135 Microtome (Leica, Germany). After assembly onto a glass slide, the tissue was deparaffinized in xylene, rehydrated in graded ethanol, stained with hematoxylin and eosin (H&E), and sealed with Entellan (Merck Millipore, Germany). The H&E slides were examined using a light microscope (Primostar, Zeiss, US). The total number of superficial follicles in the right and left ovaries was calculated manually using a light stereomicroscope (Olympus, Japan). Follicle sizes were measured using image analysis methods in ImageJ (NIH, US). Statistical analysisThe research data were analyzed using one-way analysis of variance (ANOVA). If the ANOVA test results were significantly different (p < 0.05), they were further analyzed using the Tukey test at a 95% confidence interval. ResultsWeight before and during the pregnancyTables 2 and 3 present data on the body weight of the mated and gestational mothers. The data showed no significant difference (p > 0.05). The body weight data before the mated rats in the pre-pregnancy period showed no significant difference (p > 0.05). The order of increase in body weight of rats from highest to lowest was as follows: P4 experienced an increase of 34g (19.2%), P2 by 33g (15.8%), P6 by 31g (14.02%), P1 by 29g (14.8%), P5 by 21g (11.9%), and P7 by 20g (10.3%). Rats in the P4 treatment tended to experience higher body weight gain compared with those in other treatments. Table 2. Effect of Harpodon nehereus and Chlorella vulgaris feed supplementation doses on the average body weight of pre-pregnant rats.
Table 3. Effect of Harpodon nehereus and Chlorella vulgaris feed supplementation on the average body weight of pregnant rats.
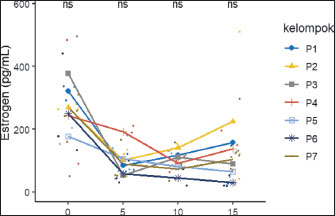
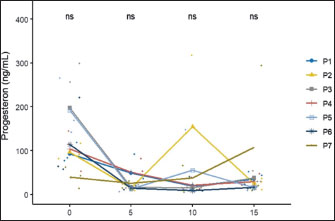
The body weight data at gestational age showed no significant difference (p > 0.05), but the P5 group tended to experience greater weight gain than the other groups. The data show no significant difference (p > 0.05); however, as the table indicates, rats in the P5 treatment tended to experience greater weight gain than rats in the other treatments. Serum estrogen and progesterone levelsThe effects of feeding H. nehereus meal and C. vulgaris powder on serum estrogen and progesterone concentrations of maternal experimental rats were observed on the day before mating (D-0) and continued at 5 days (D-5), 10 days (D-10), and 15 days (D-15) of pregnancy. Serum estrogen concentration analysis showed no significant difference (p > 0.05) between the control maternal rats (P1) and supplemented maternal rats (P2, P3, P4, P5, P6, and P7) one day before mating (D-0) and on D-5, D-10, and D-15 of pregnancy (Fig. 1). Serum estrogen concentrations 1 day before mating were numerically higher and decreased on D-5 and did not change during pregnancy (Fig. 1).
Fig. 1. Effects of H. nehereus and C. vulgaris flour supplementation on estrogen levels in experimental rats D-0=before mating; D-5=5th day of pregnancy; D-10=10th day of pregnancy, and D-15=15th day of pregnancy. P1=control; P2=pregnant mother’s milk 0.72 g/kg; P3=H. nehereus meal at a dose of 0.8 g/kg; P4=H. nehereus meal at a dose of 1.6 g/kg; P5=H. nehereus meal at a dose of 2.4 g/kg; P6=C. vulgaris powder at a dose of 0.5 g/kg; and P7= H. nehereus meal at a dose of 2.4 g/kg of feed + C. vulgaris powder at a dose of 0.5 g/kg. The patterns of serum progesterone concentrations before mating and during pregnancy were similar to those of serum estradiol concentrations. Serum progesterone concentration analysis showed no significant difference (p > 0.05) between the control maternal rats (P1) and supplemented maternal rats (P2, P3, P4, P5, P6, and P7) one day before mating (D-0) and on D-5, D-10, and D-15 of pregnancy (Fig. 2). Similar to the pattern of serum estrogen concentrations, serum progesterone concentrations one day before mating were numerically higher and decreased on D-5 and did not change during days 10 and 15 of pregnancy (Fig. 2).
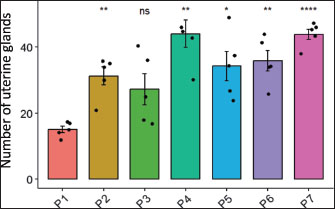
Fig. 2. Effects of H. nehereus and C. vulgaris flour supplementation on progesterone hormone levels in experimental maternal rats D-0=before mating; D-5=5th day of pregnancy; D-10=10th day of pregnancy, and D-15=15th day of pregnancy. P1=control; P2=pregnant mother’s milk 0.72 g/kg; P3=H. nehereus meal at a dose of 0.8 g/kg; P4=H. nehereus meal at a dose of 1.6 g/kg; P5=H. nehereus meal at a dose of 2.4 g/kg; P6=C. vulgaris powder at a dose of 0.5 g/kg; and P7= H. nehereus meal at a dose of 2.4 g/kg of feed + C. vulgaris powder at a dose of 0.5 g/kg. Uterine analysisThe results of the uterine analysis showed that in maternal experimental rats supplemented with pregnant milk at a dose of 0.72 g/kg feed (P2), H. nehereus meal at a dose of 1.6 g/kg feed (P4), H. nehereus at a dose of 2.4 g/kg feed (P5), C. vulgaris at a dose of 0.5 g/kg feed (P6), and combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7), there were significant increases (p < 0.05) in the number of uterine glands (corpus luteum) compared with the control group (P1) (Fig. 3). However, in the experimental maternal rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P3), there was no significant difference in the number of uterine glands compared with that in the control experimental rats (P1). The highest number of uterine glands was found in the experimental maternal rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4), followed by experimental rats in groups P7, P6, P5, P2, P3, and P1. This sequence shows that supplementation of experimental maternal rats with H. nehereus at a dose of 1.6 g/kg feed (P4) is the most influential dose in increasing uterine gland growth, and the lowest uterine gland growth was found in control experimental rats without supplementation (P1) and experimental rats supplemented with H. nehereus at the lowest dose (0.8 g/kg feed or P3).
Fig. 3. Effects of H. nehereus and C. vulgaris flours on the number of uterine glands in experimental maternal rats Statistical analysis was performed using analysis of variance and Tukey’s test. Significant differences were considered significant at ns p > 0.05; *=p < 0.05; **=p < 0.001; and ***=p < 0.0001. The results of the analysis of uterine muscle thickness showed no significant difference (p > 0.05) between the control (P1) and the other groups (P2, P3, P4, P5, P6, and P7) (Fig. 4). The muscle thickness in all treatments was in the same range (0.12–0.15) and was not significantly different. These results show that the administration of H. nehereus and C. vulgaris meals had no significant effect on increasing the endometrial layer thickness in female white rats. The proliferation and differentiation of the endometrial glands contribute to the increased thickness of the endometrial layer.
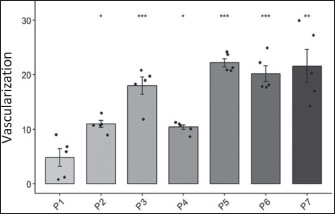
Fig. 4. Effects of H. nehereus and C. vulgaris flours on uterine muscle thickness in experimental maternal rats Statistical analysis was performed using analysis of variance and Tukey’s test. Significant differences were considered significant at ns p > 0.05; *=p < 0.05; **=p < 0.001; and ***=p < 0.0001. The results of the vascularization analysis showed a significant difference (p < 0.05) between the control group (P1) and the other treatment groups (P2, P3, P5, P6, and P7) (Fig. 5). In all groups, there was an increase in the vascularization in the experimental maternal rats, except in the experimental maternal rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4), which showed no significant difference when compared with experimental rats without supplementation (P1 or control group). The highest degree of vascularization was found in the experimental maternal rats supplemented with the highest dose of H. nehereus (2.4 g/kg feed or P5 group), followed by experimental maternal rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7), experimental maternal rats supplemented with C. vulgaris at a dose of 0.5 g/kg feed (P6), experimental maternal rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P3), experimental maternal rats supplemented with pregnant milk at a dose of 0.72 g/kg feed (P2), and experimental maternal rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4), and the lowest degree of vascularization was found in the control experimental maternal rats without supplementation (P1).
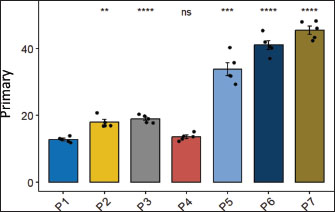
Fig. 5. Effects of H. nehereus and C. vulgaris flours on the number of vascularizations in experimental maternal rats Statistical analysis using analysis of variance and Tukey’s t-test. Significant difference with is p > 0.05; *=p < 0.05; **=p < 0.001; and ***=p < 0.0001. Ovarian follicle numberThe numbers of primary ovarian follicles in the experimental rats were significantly different (p < 0.05) between the control experimental rats without supplementation (P1) and experimental rats supplemented with pregnant milk at a dose of 0.72 g/kg feed (P2), experimental rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P3), experimental rats supplemented with H. nehereus at a dose of 2.4 g/kg feed (P5), experimental rats supplemented with C. vulgaris at a dose of 0.5 g/kg feed (P6), and experimental rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7) (Fig. 6). The higher the protein treatment dose, the higher the number of follicles in the experimental rats. However, in the experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4), there was no significant difference in the number of follicles (p > 0.05) compared with the control group.
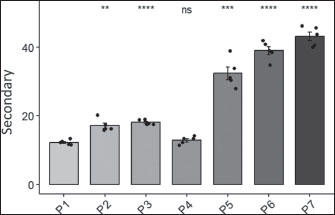
Fig. 6. Effects of H. nehereus and C. vulgaris flour supplementation on the number of primary follicles in maternal experimental rats Statistical analysis was performed using analysis of variance and Tukey’s test. Significant differences were considered significant at ns p > 0.05; *=p < 0.05; **=p < 0.001; and ***=p < 0.0001. Experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4) showed decreased primary follicles. Experimental rats supplemented with H. nehereus at a dose of 2.4 g/kg feed (P5), experimental rats supplemented with C. vulgaris at a dose of 0.5 g/kg feed (P6), and experimental rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7) showed the highest number of primary follicles compared to experimental rats supplemented with pregnant milk at a dose of 0.72 g/kg feed (P2), experimental rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P3), and experimental rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P4). In experimental rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7), it was found that the combination of H. nehereus and C. vulgaris was more effective than the combination of H. nehereus and C. vulgaris. Supplementation with Vulgaris has the highest number of secondary follicles. This demonstrates that the combination of the two ingredients positively affects the development of primary follicles and enhances the reproductive performance of the female experimental rats. The number of secondary follicles was significantly different between the control and treatment groups (p < 0.05). However, the number of secondary follicles was not significantly different between the experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4) and the control group (P1) (Fig. 7). From the data obtained, the higher the treatment dose, the higher the number of secondary follicles. From the data obtained, the higher the dose of treatment protein, the higher the number of follicles in the experimental rats. However, the number of primary follicles decreased in experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4). Experimental rats supplemented with H. nehereus at a dose of 2.4 g/kg feed (P5), experimental rats supplemented with C. vulgaris at a dose of 0.5 g/kg feed (P6), and experimental rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7) showed the highest number of follicles compared to the control experimental rats without supplementation (P1), experimental rats supplemented with pregnant milk at a dose of 0.72 g/kg feed (P2), experimental rats supplemented with H. nehereus at a dose of 0.8 g/kg feed (P3), and experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4). However, in all treatments, the number of secondary follicles was higher than that in the control group, except in experimental rats supplemented with H. nehereus at a dose of 1.6 g/kg feed (P4), which had a lower number of secondary follicles than the control. In the experimental rats supplemented with a combination of H. nehereus at a dose of 2.4 g/kg feed and C. vulgaris at a dose of 0.5 g/kg feed (P7), the combination treatment produced the highest number of secondary follicles.
Fig. 7. Effects of H. nehereus and C. vulgaris flour supplementation on the number of secondary follicles in maternal experimental rats Statistical analysis using analysis of variance and Tukey’s t-test. Significant difference with is p > 0.05; *=p < 0.05; **=p < 0.001; and ***=p < 0.0001. DiscussionIn this study, the administration of H. nehereus and C. vulgaris to adult female Sprague-Dawley rats in the diestrus phase of the estrus cycle for 20 days before mating, during pregnancy, and lactation was observed to have a positive effect on health, gonadal growth, female animal development, and offspring intelligence. Optimal ovarian growth and development are associated with the production of high-quality follicles that secrete high concentrations of estradiol and progesterone (Chauvin et al., 2022; Desta, 2024). These hormones stimulate the growth and development of the uterus and placenta, thereby creating an ideal environment for embryo and fetus growth and development during pregnancy (Tsakiridis et al., 2020; Habibi et al., 2022; Parisi et al., 2023). An optimal environment during prenatal development promotes enhanced gene set expression and improves postnatal offspring survival. Consequently, H. nehereus and C. vulgaris can improve reproductive performance. In this study, the stages of gonad development and estrus cycles of the experimental rats were different because the estrus cycles were not synchronized. Treatments were administered for 20 days before mating or for approximately four consecutive estrus cycles. Estrogen and progesterone hormone parameters were evaluated in unmated rats after 20 days of treatment. Uterine and ovarian parameters were assessed immediately after delivery. The gestational age body weight data showed no significant difference (p > 0.05), but the P5 group tended to experience more weight gain than the other groups. The order of body weight of the experimental rats was from highest to lowest in sequence, as follows: P5, P4, P1, P2, P6, P7, and P3. This suggests that providing a mixed feed of H. nehereus and C. vulgaris can influence the increased body weight of the treated rats during pregnancy. Protein consumption during pregnancy significantly affects the development and health of the mother and fetus (Herring et al., 2018; Castro-Rodríguez et al., 2020). Research indicates that the balance between protein and energy in the diet is crucial for supporting optimal growth and development. Hutasoit et al. (2021) found that the right balance of protein and total digestible nutrients can increase BWG and feed conversion efficiency in pregnant animals. This study emphasizes the importance of a balanced diet to support animal reproductive health. Adequate and balanced protein consumption during pregnancy in pregnant rats supports fetal growth and affects maternal health. These studies suggest that a diet rich in protein and other essential nutrients is crucial for ensuring optimal health and healthy development in offspring. Overall, a nutritional combination that includes high-quality protein, omega-3 and omega-6 fatty acids, and flavonoids, which are crucial during the reproductive phase, will provide metabolic support, promote growth, and protect against oxidative stress. The results of this study showed the potential of H. nehereus and C. vulgaris supplementation to improve health, ovarian and uterine growth, increase hormone levels, and promote the growth and development of female animals. Optimal ovarian growth and development will produce high-quality follicles, positively impacting the production of high amounts of estrogen (E2) and progesterone hormones (Herring et al., 2018; Sood et al., 2021; Pan and Huang, 2022). Estrogen and progesterone are hormones that stimulate the growth and development of the uterus and placenta to prepare and provide an ideal environment to support the growth and development of the embryo and fetus during pregnancy (Panda et al., 2021; Tal and Taylor, 2021; Chauvin et al., 2022; Dukic et al., 2024). An optimal environment during prenatal development leads to better gene expression, resulting in healthier and superior offspring (Enquobahrie et al., 2022; Socha et al., 2024). Serum estrogen levels showed no significant difference (p > 0.05) across all treatments; however, there was a tendency for an increase in the treatment group compared to the control. The hormone estrogen produced by the developing follicles before ovulation during estrus and progesterone produced by the corpus luteum and placenta during pregnancy initiate the reproductive process in female mammals (Manalu, 1999). These pregnancy hormones are the initial signals that initiate a comprehensive reproductive process, starting from the preparation of the ovum and sperm, fertilization, preparation of the uterus for implantation, and development of the placenta, embryo, and fetus (Gad et al., 2017; Sferruzzi-Perri et al., 2017; Savirón-Cornudella and Herraiz, 2022; Aguilera et al., 2022; Kodithuwakku et al., 2024), which ultimately affect the flow of nutrients to the developing embryo and fetus. Progesterone hormone levels from the treatment showed no significant difference (p > 0.05) between the treatment groups from day 0 to day 10 of pregnancy. However, a substantial difference was observed between the groups and the control group on day 15. During pregnancy, progesterone hormone levels in the experimental rats undergo significant changes, which are essential in maintaining pregnancy and preparing the body for labor. Progesterone, which is produced mainly by the corpus luteum and then by the placenta, prepares the endometrium for embryo implantation and maintains pregnancy by inducing a calm state in the uterus. The development of primary and secondary follicles in the experimental rats was significantly different, with the treatment group having more follicles than the control group. Supplementation of H. nehereus and C. vulgaris influences the development of primary and secondary follicles. Sufficient protein levels in the feed also regulate the secretion of follicle-stimulating hormones and luteinizing hormone (FSH and LH), which are essential for follicle development. FSH and LH are glycoprotein molecules that stimulate follicle growth and ovulation (Herring, 2018; Lin et al., 2025). The number of corpora luteas describes the number of successfully ovulated ova. The number of corpora luteas in the treatment group showed significantly different results, with the treatment group having a higher number of corpora luteas than the control group. Providing fish meal and C. vulgaris powder increased the number of mature eggs, as indicated by a higher number of corpora luteas compared with the control. The greater the number of corpora lutea, the higher the success rate of ovulation in the maternal experimental rats. Supplementing experimental female rats with H. nehereus and C. vulgaris powder can improve reproductive performance. Vascularization in the treated experimental rats showed a significant effect. The high protein content in the ration provides essential amino acids crucial for forming structural proteins and enzymes necessary for angiogenesis (the formation of new blood vessels) (Gou et al., 2017; Li et al., 2022). During pregnancy, the uterus requires a greater blood supply, and adequate protein provision supports the formation of blood vessel tissue to meet the fetus’s oxygen and nutrient needs (McLean et al., 2017; Man et al., 2023). Omega-3 from fish powder plays an important role in supporting blood vessel health and stimulates the production of prostaglandins that function in healthy inflammatory processes and support blood vessel formation. Omega-3 fatty acids increase blood flow and vessel flexibility, strengthening the uterine vascular system and supporting embryo growth and development during pregnancy (Drenjančević and Pitha, 2022; Dos Santos et al., 2024). The combination of flavonoids, proteins, omega-3, omega-6, and omega-9 can synergistically support vascularization. Flavonoids act as antioxidants and increase blood vessel growth factors, proteins provide raw materials for tissue synthesis, and omega-3 supports blood vessel function. These nutrients help maximize blood vessel formation in the uterus, which is important for meeting the fetus’s oxygen and nutrient needs (Dong et al., 2022; Wang et al., 2022; Zhao et al., 2023; Li et al., 2023). Flavonoids in C. vulgaris are potent antioxidants that help reduce uterine oxidative stress. Low oxidative stress promotes a healthy environment for angiogenesis (new blood vessel formation). Flavonoids can also increase the production of blood vessel growth factors, such as VEGF, which plays a crucial role in triggering and supporting the formation of new blood vessels in the uterus (Ciumărnean et al., 2020; Khan et al., 2021). The combination of protein, omega-3 fatty acids, flavonoids, and nutrients from C. vulgaris can enhance the response of the uterus to progesterone, which is crucial in preparing the uterine lining for implantation and pregnancy. Progesterone also plays a role in stimulating blood vessel formation in the endometrium; therefore, increasing nutrient intake can enhance the effect of progesterone on stimulating uterine angiogenesis. During pregnancy, the uterus must provide adequate oxygen and nutrients to support fetal growth (Marshal et al., 2022). Increasing nutrient intake that supports angiogenesis helps the body meet these needs by increasing vascularization (Kissane and Egginton, 2019; Akbarian et al., 2022). Blood vessels in the uterus are stimulated with the supply of protein, unsaturated fatty acids, and C. vulgaris, allowing optimal blood flow to the embryo. The combination of high protein, omega-3 fatty acids, and antioxidant nutrients synergistically supports angiogenesis. Protein provides the building blocks; omega-3 fatty acids strengthen blood vessel function and flexibility, while C. vulgaris antioxidants help maintain a healthy uterine environment for optimal blood vessel development. The nutrients from the high protein, omega-3s, and flavonoids in C. vulgaris support and stimulate uterine vascularization. An increased number of healthy blood vessels is essential to ensure adequate blood supply to the fetus and support optimal development during pregnancy. Providing feed supplementation with H. nehereus and C. vulgaris can be a recommendation to improve the mother’s performance, ultimately affecting the offspring’s quality. ConclusionSupplementation of experimental rats with H. nehereus and C. vulgaris fishmeal resulted in elevated estrogen and progesterone levels, an increase in both primary and secondary follicles, and the development of uterine glands. Moreover, maternal blood vessel formation was enhanced following supplementation therapy. These results suggest that adding fishmeal and C. vulgaris to female rats’ diets can enhance reproductive performance, influencing pre-pregnancy and pregnancy body weight, estrogen and progesterone levels, uterine gland quantity, uterine muscle thickness, degree of vascularization, primary follicle count, and secondary follicle count. AcknowledgmentsThe authors express their gratitude to the Center for Education Financial Services, Beasiswa Pendidikan Indonesia (BPI), the Ministry of Education of the Republic of Indonesia, the Indonesia Endowment Fund for Education (LPDP), and the Ministry of Finance of the Republic of Indonesia for their generous financial support. Author’s contributionsAll authors have contributed to the final version of the manuscript. The authors’ contributions are as follows: NA proposed the idea, collected, analyzed, and interpreted the data, and drafted the original manuscript. WM, IKMA, and AA conceptualized the idea, refined the research methodology, interpreted the data, and contributed to writing, review, and editing. All authors have read, reviewed, and approved the final version of the manuscript. Ethical approvalThe Animal Ethics Committee of the School of Veterinary Medicine and Biomedical Sciences, Bogor Agricultural University, reviewed and approved all experimental procedures. The number is 107/KEH/SKE/IX/2023. Animal treatment was conducted with minimum pain or discomfort, following the guidelines established by the Institutional Animal Ethics Committee. The ethics approval was granted on September 7, 2023. Conflict of interestAll authors declare no conflicts of interest. FundingThis research was supported by a grant from the Center for Education Financial Services, Beasiswa Pendidikan Indonesia (BPI) 2022, a collaboration between the Ministry of Education of the Republic of Indonesia and the Indonesia Endowment Fund for Education (LPDP), in collaboration with the Ministry of Finance of the Republic of Indonesia. Data availabilityAll data are available in the revised manuscript. ReferencesAguilera, N., Salas-Pérez, F., Ortíz, M., Álvarez, D., Echiburú, B. and Maliqueo, M. 2022. Rodent models in placental research: implications for fetal origins of adult disease. Anim. Reprod. 19(1), e20210134; doi:10.1590/1984-3143-AR2021-0134 Akbarian, M., Bertassoni, L.E. and Tayebi, L. 2022. Biological aspects in controlling angiogenesis: current progress. Cell. Mol. Life Sci. 79, 349; doi:10.1007/s00018-022-04348-5 Andriyanto, A., Widi, L.N., Subangkit, M., Tarigan, E., Irarang, Y., Nengsih, R.F. and Manalu, W. 2022. Potential use of maceration of Indonesian basil (Ocimum basilicum) to increase synthesis and secretion of estradiol and progesterone to improve the prenatal growth of offspring using female albino rats as an animal model. Vet. World 15(5), 1197–1207; doi:10.14202/vetworld.2022.1197-1207 Castro-Rodríguez, D.C., Rodríguez-González, G.L., Menjivar, M. and Zambrano, E. 2020. Maternal interventions to prevent adverse fetal programming outcomes from maternal malnutrition: evidence from animal models. Placenta 102, 49–54; doi:10.1016/j.placenta.2020.04.002 Chauvin, S., Cohen-Tannoudji, J. and Guigon, C.J. 2022. Estradiol signaling at the heart of folliculogenesis: its potential deregulation in human ovarian pathologies. Int. J. Mol. Sci. 23(1), 512; doi:10.3390/ijms23010512 Ciumărnean, L., Milaciu, M.V., Runcan, O., Vesa, S.C., Răchișan, A.L., Negrean, V., Perné, M.G., Donca, V.I., Alexescu, T.G., Para, I. and Dogaru, G. 2020. The effects of flavonoids in cardiovascular diseases. Molecules 25(18), 4320; doi:10.3390/molecules25184320 Desta, A.G. 2024. Effect of crude protein and energy on dairy cow conception: a review. Discov. Anim. 1, 29; doi:10.1007/s44338-024-00030-1 Dong, J., Wang, Y. and Zhang, J. 2022. Maternal nutrient intake during pregnancy affects fetal development and maternal health in rats: a systematic review. Nutrients 14(11), 1–13; doi:10.3390/nu14112471 Dos Santos, S.P., Kra, G. and Butenko, Y. 2024. Maternal supplementation with n-3 fatty acids affects placental lipid metabolism, inflammation, oxidative stress, endocannabinoid system, and neonate cytokine concentrations in dairy cows. J. Anim. Sci. Biotechnol. 15, 74; doi:10.1186/s40104-024-01033-4 Drenjančević, I. and Pitha, J. 2022. Omega-3 polyunsaturated fatty acids—vascular and cardiac effects on the cellular and molecular level (narrative review). Int. J. Mol. Sci. 23(4), 69–79; doi:10.3390/ijms23042104 Dukic, J., Johann, A., Henninger, M. and Ehlert, U. 2024. Estradiol and progesterone levels from pregnancy to postpartum: a longitudinal latent class analysis. Front. Glob. Womens. Health 5, 1428494; doi:10.3389/fgwh.2024.1428494 Enquobahrie, D.A., MacDonald, J., Hussey, M., Bammler, T.K., Loftus, C.T., Paquette, A.G., Byington, N., Marsit, C.J., Szpiro, A., Kaufman, J.D., LeWinn, K.Z., Bush, N.R., Tylavsky, F., Karr, C.J. and Sathyanarayana, S. 2022. Prenatal exposure to PM and placental gene expression. Environ. Int. 165, 107310; doi:10.1016/j.envint.2022.107310 Gad, A., Lachman, M. and Koren, G. 2017. Placental transfer of maternal insulin-like growth factor-I and insulin-like growth factor-binding protein-3 in early human pregnancy. J. Obstet. Gynaecol. Res. 37(1), 1–8; doi:10.1111/j.1447-0756.2010.01385.x Gaskins, A.J., Chavarro, J.E. and Hauser, R. 2016. Dietary patterns and semen quality in men attending a fertility clinic. Hum. Reprod. 31(5), 976–984; doi:10.1093/humrep/dew050 Guo, D., Murdoch, C.E., Xu, H., Shi, H., Duan, D.D., Ahmed, A. and Gu, Y. 2017. Vascular endothelial growth factor signaling requires glycine to promote angiogenesis. Sci. Rep. 7, 14749; doi:10.1038/s41598-017-15246-3 Habibi, N., Mousa, A., Tay, C.T., Khomami, M.B., Patten, R.K., Andraweera, P.H., Wassie, M., Vandersluys, J., Aflatounian, A., Bianco-Miotto, T., Zhou, S.J. and Grieger, J.A. 2022. Maternal metabolic factors and the association with gestational diabetes: a systematic review and meta-analysis. Diabetes Metab. Res. Rev. 38(5), 3532; doi:10.1002/dmrr.3532 Herring, C.M., Bazer, F.W., Johnson, G.A. and Wu, G. 2018. Maternal dietary protein intake affects fetal survival, growth, and development. Exp. Biol. Med. 243(6), 525–533; doi:10.1177/1535370218758275 Hutasoit, R., Solehudin, S., Petrus Ginting, S., Simanihuruk, K., Zubaidah, S. and Sumarni, S. 2021. Dietary crude protein and total digestible nutrient effects on the growth phase performance of Boerka goats. Int. J. Trop. Vet. Biomed. Res. 6(1), 18–23; doi:10.21157/ijtvbr.v6i1.23869 Jiang, W., Zhao, Y., Wu, X., Du, Y. and Zhou, W. 2023. Global protein-energy malnutrition health inequalities from 1990 to 2019 and forecast prevalence for 2044: data from the Global Burden of Disease Study 2019. Public Health 225, 102–109; doi:10.1016/j.puhe.2023.10.003 Khan, J., Deb, P.K., Priya, S., Medina, K.D., Devi, R., Walode, S.G. and Rudrapal, M. 2021. Dietary flavonoids: cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules 26(13), 4021; doi:10.3390/molecules26134021 Kissane, R.W. and Egginton, S. 2019. Exercise-mediated angiogenesis. Curr. Opin. Physiol. 10, 193–201; doi:10.1016/j.cophys.2019.06.005 Ko, M.J., Mulia, G.E. and Van Rijn, R.M. 2019. Common anesthesia/euthanasia methods for brain collection differentially impact MAPK activity in male and female C57BL/6 rats. Front. Cell. Neurosci. 13, 96; doi:10.3389/fncel.2019.00096 Kodithuwakku, S.P., Ng, E.H.Y., Yeung, W.S.B., Lee, K.F. and Zhu, Y.Q. 2024. An update on hormonal regulation of early embryo-fallopian tube interactions: insights for improving outcomes of assisted reproductive technology. Reprod. Dev. Med. 8(1), 39–49; doi:10.1097/RD9.0000000000000074 Li, J., Liu, X. and Chen, Y. 2023. Nutrient intake during pregnancy affects fetal development and maternal health in rats: a systematic review. J. Dev. Orig. Health Dis. 14(2), 153–164; doi:10.1017/S204017442200039X Li, M., Wu, Y. and Ye, L. 2022. The role of amino acids in endothelial biology and function. Cells 11(8), 1372; doi:10.3390/cells11081372 Likhar, A. and Patil, M.S. 2023. Importance of maternal nutrition in the first 1,000 days of life and its effects on child development: a narrative review. Cureus 14(10), e30083; doi:10.7759/cureus.30083 Lin, L.T., Li, C.J., Lee, Y.S. and Tsui, K.H. 2025. Recombinant follicle-stimulating hormone and luteinizing hormone enhance mitochondrial function and metabolism in aging female reproductive cells. Int. J. Mol. Sci. 26(1), 83; doi:10.3390/ijms26010083 Man, A.W.C., Zhou, Y., Xia, N. and Li, H. 2023. Dietary supplements and vascular function in hypertensive disorders of pregnancy. Pflugers. Arch. - Eur. J. Physiol. 475(7), 889–905; doi:10.1007/s00424-023-02810-2 Manalu, W., Sumaryadi, M.Y., Sudjatmogo. and Satyaningtijas, A.S. 2000. Effect of superovulation prior to mating on milk production performance during lactation in ewes. J. Dairy Sci. 83(3), 477–483; doi:10.3168/jds.S0022-0302(00)74906-X Manalu, W. 1999. Correlations of serum progesterone concentration with uterine and fetal weights at weeks 7 and 15 of pregnancy in thin-tail ewes in Java. Anim. Biosci. 12(6), 854–861; doi:10.5713/ajas.1998.300 Marshall, N.E., Abrams, B., Barbour, L.A., Catalano, P., Christian, P., Friedman, J.E., Hay, W.W., Hernandez, T.L., Krebs, N.F., Oken, E., Purnell, J.Q., Roberts, J.M., Soltani, H., Wallace, J. and Thornburg, K.L. 2022. The importance of nutrition in pregnancy and lactation: lifelong consequences. Am. J. Obstet. Gynecol. 226(5), 607–632; doi:10.1016/j.ajog.2021.12.035 McLean, K.J., Crouse, M.S., Crosswhite, M.R., Pereira, N.N., Dahlen, C.R., Borowicz, P.P., Reynolds, L.P., Ward, A.K., Neville, B.W. and Caton, J.S. 2017. Impacts of maternal nutrition on uterine and placental vascularity and mRNA expression of angiogenic factors during the establishment of pregnancy in beef heifers. Transl. Anim. Sci. 1(2), 160–167; doi:10.2527/tas2017.0019 National Research Council. 2011. Guide for the care and use of laboratory animals. 8th edition. Washington, DC: The National Academies Press. Okada, H., Tsuzuki, T. and Murata, H. 2018. Decidualization of the human endometrium. Repr. Med. Biol. 17(3), 220–227; doi:10.1002/rmb2.12088 Paliwal, C. and Jutur, P.P. 2021. Dynamic allocation of carbon flux triggered by task-specific chemicals is an effective, non-gene-disruptive strategy for sustainable and cost-effective algal biorefineries. Chem. Eng. J. 418, 129413; doi:10.1016/j.cej.2021.129413 Pan, P. and Huang, X. 2022. Clinical application of growth hormone and its biological and molecular mechanisms in assisted reproduction: a review. Int. J. Mol. Sci. 23(18), 10768; doi:10.3390/ijms231810768 Panda, S., Das, A., Mallik, A. and Baruah, S. 2021. Normal puerperium. In Empowering midwives and obstetric nurses. Ed. Ray, A. London, United Kingdom: IntechOpen; doi:org/10.5772/intechopen.96348 Parisi, F., Fenizia, C., Introini, A., Zavatta, A., Scaccabarozzi, C., Biasin, M. and Savasi, V. 2023. The pathophysiological role of estrogens in the initial stages of pregnancy: molecular mechanisms and clinical implications for pregnancy outcome from the periconceptional period to the end of the first trimester. Hum. Reprod. Update 29(6), 699–720. Salim, G., Rukisah, R., Azis, T., Tamrin, A., Awaludin, P., Nugraeni, C., Al-Hafidz, M., Rahmatsyah, M., Herry, M., Muslikah, V., Safira, R., Sastri, S., Selly, S., Jati, T., Jalil, A., Fauzi, M., Lailaturrif’ah, M. and Wulandari, E. 2022. Processing of nomei fish (H. nehereus) into nuggets in the marine region of Tarakan City. Jompa Abdi 1(1), 126–136. Sathasivam, R., Radhakrishnan, R., Hashem, A. and Abdulallah, E.F. 2019. Microalgae metabolites: a rich source of food and medicine. Saudi J. Biol. Sci. 26(4), 709–722; doi:10.1016/j.sjbs.2017.11.003 Savirón-Cornudella, R. and Herraiz, I. 2022. Hormones and growth factors. Gynecol. Endocrinol. 38(3), 191–192; doi:10.1080/09513590.2022.2044469 Sferruzzi-Perri, A.N., Sandovici, M., Constancia. and Fowden. 2017. Placental phenotype and the insulin-like growth factors: resource allocation to fetal growth. J. Physiol. 595(15), 5057–5093; doi:10.1113/JP273330 Siddiqa, M., Zubair, A., Kamal, A., Ijaz, M. and Aljeddani, S.M.A. 2024. Risk factors of child malnutrition under 5 years: evidence from Pakistan using the composite index of anthropometric failure. Nutrition 127, 112523; doi:10.1016/j.nut.2024.112523 Socha, M.W., Flis, W. and Wartęga, M. 2024. Epigenetic genome modifications during pregnancy: the impact of essential nutritional supplements on DNA methylation. Nutrients 16(5), 678; doi:10.3390/nu16050678 Sood, A., Mohiyiddeen, G., Ahmad, G., Fitzgerald, C., Watson, A. and Mohiyiddeen, L. 2021. Growth hormone for in vitro fertilization. Cochrane Database Syst. Rev. 11, CD000099; doi:10.1002/14651858.CD000099.pub4 Tal, R. and Taylor, H.S. 2021. Pregnancy endocrinology. In Endotext [Internet]. Eds., Feingold, K.R., Ahmed, S.F., Anawalt, B. et al. South Dartmouth, MA: MDText.com, Inc. Available via https://www.ncbi.nlm.nih.gov/books/NBK278962/ Tsakiridis, I., Kasapidou, E., Dagklis, T., Leonida, I., Leonida, C., Bakaloudi, D.R. and Chourdakis, M. 2020. Nutrition in pregnancy: a comparative review of major guidelines. Nutrients 12(11), 3019; doi:10.3390/nu12113019 Wang, C.A., Onyeaka, H., Miri, T. and Soltani, F. 2024. Chlorella vulgaris as a food substitute: applications and benefits in the food industry. J. Food Sci. 89(12), 8231–8247; doi:10.1111/1750-3841.17529 Wang, J., Wu, J., Zhang, Y. and Zhang, J. 2022. Nutrient supplementation during pregnancy improves fetal growth and maternal health. Am. J. Clin. Nutr, 115(3), 744–754. doi:10.1093/ajcn/nqab387 World Health Organization. 2021. World health statistics 2021: monitoring health for the SDGs, Sustainable Development Goals. Geneva: World Health Organization. Zhao, Y., Zhang, J. and Li, M. 2023. Effects of maternal nutrient intake during pregnancy on fetal development and maternal health in rats: a systematic review. J. Agric. Food Chem. 71(2), 553–563; doi:10.1021/acs.Java.2c0629 | ||

| How to Cite this Article |
| Pubmed Style Nurasmi N, Manalu W, Adnyane IKM, Andriyanto A. Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Vet. J.. 2025; 15(9): 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 Web Style Nurasmi N, Manalu W, Adnyane IKM, Andriyanto A. Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. https://www.openveterinaryjournal.com/?mno=254528 [Access: June 22, 2026]. doi:10.5455/OVJ.2025.v15.i9.38 AMA (American Medical Association) Style Nurasmi N, Manalu W, Adnyane IKM, Andriyanto A. Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Vet. J.. 2025; 15(9): 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 Vancouver/ICMJE Style Nurasmi N, Manalu W, Adnyane IKM, Andriyanto A. Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Vet. J.. (2025), [cited June 22, 2026]; 15(9): 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 Harvard Style Nurasmi, N., Manalu, . W., Adnyane, . I. K. M. & Andriyanto, . A. (2025) Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Vet. J., 15 (9), 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 Turabian Style Nurasmi, Nurasmi, Wasmen Manalu, I Ketut Mudite Adnyane, and Andriyanto Andriyanto. 2025. Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Veterinary Journal, 15 (9), 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 Chicago Style Nurasmi, Nurasmi, Wasmen Manalu, I Ketut Mudite Adnyane, and Andriyanto Andriyanto. "Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats." Open Veterinary Journal 15 (2025), 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 MLA (The Modern Language Association) Style Nurasmi, Nurasmi, Wasmen Manalu, I Ketut Mudite Adnyane, and Andriyanto Andriyanto. "Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats." Open Veterinary Journal 15.9 (2025), 4310-4321. Print. doi:10.5455/OVJ.2025.v15.i9.38 APA (American Psychological Association) Style Nurasmi, N., Manalu, . W., Adnyane, . I. K. M. & Andriyanto, . A. (2025) Harpodon nehereus and Chlorella vulgaris dietary supplementation as a strategy to enhance estrogen, progesterone, and reproductive outcomes in female rats. Open Veterinary Journal, 15 (9), 4310-4321. doi:10.5455/OVJ.2025.v15.i9.38 |

