




| Review Article | ||
Open Vet. J.. 2025; 15(6): 2329-2342 Open Veterinary Journal, (2025), Vol. 15(6): 2329-2342 Review Article A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A reviewMutasem Abuzahra1,2, Lucia Tri Suwanti2,3* and Muchammad Yunus31Department of Animal Production and Protection, Faculty of Agricultural Engineering, Hebron University, Hebron, Palestine 2Toxoplasma Study Group, Institute of Tropical Disease, Universitas Airlangga, Surabaya, Indonesia 3Division of Parasitology, Faculty of Veterinary Medicine, Universitas Airlangga, Surabaya, Indonesia * Corresponding Author: Lucia Tri Suwanti, Toxoplasma Study Group, Institute of Tropical Disease, Universitas Airlangga, Surabaya, Indonesia. Email: lucia-t-s [at] fkh.unair.ac.id Submitted: 09/02/2025 Revised: 05/05/2025 Accepted: 13/05/2025 Published: 30/06/2025 © 2025 Open Veterinary Journal
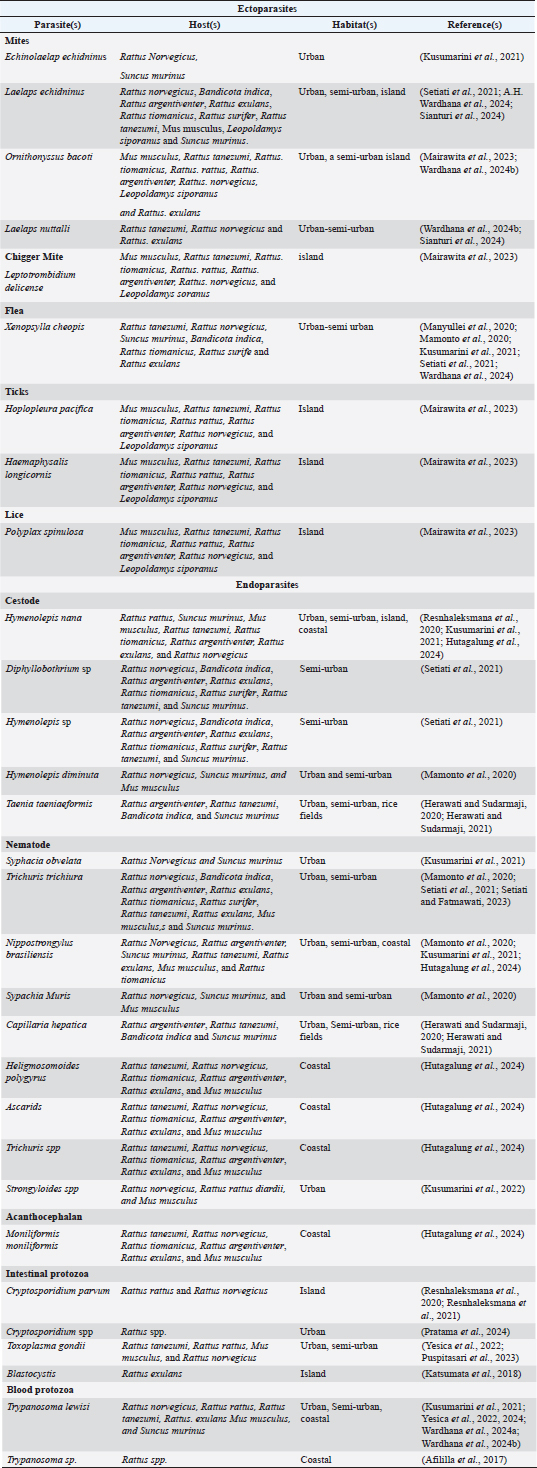
AbstractZoonotic diseases, which are transmitted between vertebrate animals and people, significantly impact global public health. Wild rats are reservoirs for numerous zoonotic illnesses, such as leptospirosis, toxoplasmosis, trypanosomiasis, and helminth infections. This study analyzed the prevalence and diversity of ectoparasites and endoparasites in wild rats in Indonesia over a decade (2015–2025). We conducted a comprehensive search using PubMed, Google Scholar, Scopus, and Science Direct, which yielded 62 publications, of which 25 met the inclusion criteria. Significant variations in ectoparasite and endoparasite loads were observed in wild rats across regions, seasons, and host species, reflecting infestation across a wide range of habitats, including urban, semi-urban, island, and rice field environments. Although molecular techniques are increasingly being applied in parasitology, only six studies used them for parasite identification in wild rats. This review highlights research gaps related to small sample sizes, limited data on blood parasites, and insufficient molecular characterization. Findings suggest that environmental changes—such as urbanization, deforestation, agricultural expansion, and seasonal shifts—may influence parasite distribution patterns. Keywords: Indonesia, wild rats, Ectoparasites, Endoparasites, parasites.. IntroductionZoonotic diseases, which are transmitted between vertebrate animals and humans, represent a significant global health challenge. Climate change and environmental degradation have intensified the emergence and re-emergence of zoonotic diseases, with approximately 60.3% of zoonoses being transmitted by domestic animals and 71.8% by wild animals (Nieto et al., 2012; Yeh et al., 2018; Behl et al., 2022). Among the major zoonotic disease reservoirs, rodents, particularly rats, play a pivotal role as carriers of pathogens such as Leptospira spp., Trypanosoma lewisi, Toxoplasma gondii, and various helminths (Widayati et al., 2020; Palomba et al., 2023; Shirvan et al., 2024). Indonesia’s 17,000-island archipelago is a zoonotic hotspot driven by dense populations, poor hygiene in wet markets, and close human–animal interactions. Wild rats thrive in urban/rural ecosystems, amplifying disease risks via their adaptability to human habitats (Kusuma et al., 2020; Setiati et al., 2021; Setiati and Fatmawati, 2023). Tropical cities face heightened rodent-related challenges: poor infrastructure, heat, and humidity enhance pathogen survival and arthropod vector proliferation (Bonell et al., 2017). Furthermore, numerous arthropod vectors of rodent-borne diseases flourish in tropical environments, such as ticks, mites, and mosquitoes, thereby heightening the risk of human sickness (Bonell et al., 2017; Lau et al., 2020). Ninety percent of future urban growth will occur in tropical/subtropical Asian and African cities, exacerbating zoonotic risks (Bergamo and Santos, 2014). The swift proliferation of metropolitan regions has exacerbated public and environmental health issues, especially those related to rats. However, studies on urban rodent ecology and related zoonoses have focused on temperate cities. Initial research in tropical regions is still limited to a few places, such as Bangkok, Kuala Lumpur, Morogoro, and Salvador, as noted in the study by Blasdell et al. (2022). Despite the increasing relevance of urban ecosystems for public health in tropical places, there is currently insufficient understanding of how rodents exploit these surroundings, what allows them to survive, and how urbanization increases zoonotic disease risks (Blasdell et al., 2022). Rodents are small, synanthropic animals that host over 143 infectious agents—14 viruses, 31 bacteria, 83 parasites, and 15 fungi. The species are dispersed globally, with 2 277 identified species (Issae et al., 2023). Rodents can spread several microbial infections, both directly and indirectly, encompassing at least 85 zoonotic pathogens. They function as definitive and intermediate hosts for ectoparasites (vector-borne illnesses) (Hardgrove et al., 2021). Ecologically, rodents are among the most significant carriers of zoonoses (Zhang et al., 2022; Wardhana et al., 2024a). A significant number of infectious diseases that affect humans have been found to have the potential to be transmitted from animals to humans (Chan et al., 2024). Zoonotic infections, particularly those associated with rats, pose significant health risks. The increasing prevalence of these species may be due to overpopulation, habitat destruction, urbanization, and human migration (Hassell et al., 2017). Urban residential zones promote closer human–wildlife interactions, increasing the risk of transmission (Chan et al., 2024). Rattus species, especially Rattus norvegicus, are among the rodents that pose the greatest threat to metropolitan areas because of their propensity for reproduction, zoonotic potential, and near closeness to people (Archer et al., 2017). These rats frequently host arthropod ectoparasites, such as lice, fleas, ticks, and mites. The most common ectoparasites are Echinolaelaps echidnius (mites) and Polyplax spinulosa (lice). Echinolaelaps echidnius spp. are spiny, coarse-haired mites, while P. spinulosa is a hematophagous louse with a robust, oval, yellow-brown body measuring 0.6–1.5 mm (Jena et al., 2017; Kusuma et al., 2020; Kang et al., 2022; Chan et al., 2024) . The study of parasite pathogens in wild rats has been increasingly important in recent years due to their significance in the transmission of diseases such as leptospirosis, toxoplasmosis, trypanosomiasis, and helminthiasis. This study covers major findings from the past decade on parasite identification in wild rats in Indonesia, examining detection methodologies, illness incidence by location, and the consequences for public and environmental health. Materials and MethodsDatabases, search strategy, and article screeningThis research examined published publications on rodent parasites in PubMed, Google Scholar, Scopus, and Science Direct. We conducted keyword searches on the relevant issue, using terms, such as “parasite,” “ectoparasite,” “endoparasite,” “Indonesia,” “wild rat,” and “zoonosis.” Duplicated or irrelevant articles were excluded from the review. A database search between 2015 and 2025 yielded sixty-two publications. The elimination of duplicates in acquired data reduced the total number of duplicates to 35. After the screening process, five articles were excluded, resulting in a total of 25 articles. Exclusion and inclusion criteriaThe inclusion criteria encompassed original research articles without methodological limits, specifically focusing on ectoparasites and endoparasites in rodents in Indonesia, provided they were published in the English language. The publishing span was 10 years (2015–2025) to fulfill the review objectives. The results of parasite investigations conducted over the past decade have remained pertinent and adequate to address the review objectives. A total of 25 articles on the specified topic were obtained and examined. Overview of parasites in Indonesian rodents: habitats and distributionThis review discusses the problems with earlier research that examined parasites (ectoparasites and endoparasites) in wild rats in Indonesia. These challenges encompass limited sample sizes, insufficient data on blood parasites, and the absence of molecular methods. Twenty-five published research studies on parasites in wild rats conducted in Indonesia during the past decade were synthesized, reviewed, and grouped according to parasite kinds in Table 1. Researchers have employed diverse methodologies and approaches to detect and identify ectoparasite and endoparasite species in wild rats. Techniques for ectoparasite identification include fur combing, direct inspection, skin scraping, culturing of immature ticks, and molecular methods such as PCR followed by DNA sequencing to detect gene polymorphisms. The techniques for endoparasite detection included blood smear analysis, necropsy, direct stool examination, immunofluorescent antibody assays, in vitro cultivation, genetic methodologies, and histopathological evaluation. In 13 rat species in Indonesia, we documented nine ectoparasites and 21 endoparasites, with approximately 1,500 examined rats. The study was conducted in a variety of Indonesian regions, with samples ranging from urban and nonurban rat species to ectoparasites and endoparasites. The majority of studies concentrated on urban and semi-urban wild rats, while few looked at non-urban wild rats, such as those found in rice fields, coastal areas, and islands. Samples were gathered from 15 different sites in Indonesia, spanning five major islands, with a primary focus on Java Island, with the majority collected during two seasonal periods: October to February and March to August. As a result, researchers have documented the parasites that infect wild rats in Indonesia. The published research in Indonesia predominantly concentrated on Java Island, the region with the highest density, whereas only a limited number of studies were undertaken in Sumatra and other regions of the country. Zoonotic ectoparasites in rodents inhabiting Indonesia: prevalence, transmission, and public health implicationsEctoparasites are parasites that live on the external surface of the host. They include ticks, fleas, mites, lice, and some insects (Ho et al., 2021; Materu et al., 2022), which can infest humans, wild animals, and domestic animals (Materu et al., 2022; Chan et al., 2024). Ectoparasites are primarily transmitted through direct contact, gradually moving to attach to and burrow into the superficial layers of the host’s skin. Once attached, they typically infest areas such as the neck, shoulders, and midbody, leading to clinical manifestations such as scratching, small wounds, dermatitis, and, in severe cases, anemia (Chan et al., 2024). Table 1. Table 1. Ectoparasites and endoparasites in wild rats in Indonesia identified in studies conducted from 2015 to 2025
Despite being overlooked, lice, fleas, hard ticks, and mites pose significant health risks due to their ability to infest a wide range of hosts, including humans. They act as vectors, weakening the health of other species by transmitting various pathogens (Materu et al., 2022; Chan et al., 2024). Ectoparasites are known to transmit zoonotic pathogens, significantly affecting public health. Over 15 ectoparasite species can be harbored by a single rodent, contaminating numerous other rodent species. This increases the pathogen mutation risk as well as zoonotic disease spreading risk (Salas et al., 2019; Ho et al., 2021). Among the most notorious ectoparasites associated with rodents, Xenopsylla cheopis (oriental rat flea), a well-known vector responsible for the spread of Yersinia pestis, the bacterium that caused the bubonic plague pandemic and urban scrub typhus (de Almeida et al., 2020). Ornithonyssus bacoti (tropical rat mite): A carrier of hantavirus infections, which transmits disease from rodents to humans (Younis et al., 2017) There have been seven published research studies on ectoparasite infections in Indonesian rodents over the past 10 years (Manyullei et al., 2020; Mamonto et al., 2020; Kusumarini et al., 2021; Setiati et al., 2021; Mairawita et al., 2023; Wardhana et al., 2024b; Sianturi et al., 2024). The studies were carried out in a variety of habitats throughout Indonesia, including cities, semiurban areas, islands, and rice fields, with a concentration in the Java region. Despite extensive research on rodent-associated parasites, a significant methodological limitation persists across many studies: the reliance on insufficient sample sizes, which compromises the accuracy of parasite prevalence and abundance estimations. Among the six studies analyzed, only one (Setiati et al., 2021) met the recommended threshold of >80 host individuals, examining 89 rodents. In contrast, most studies utilized markedly smaller sample sizes, often below the critical threshold required for robust statistical reliability, and met the recommended threshold of >80 host individuals, examining 89 rodents. In stark contrast, the majority of studies utilized markedly smaller sample sizes, often falling below the critical threshold required for robust statistical reliability. This highlights the need for larger sample sizes to reduce bias and improve the accuracy of parasitological studies. However, the aggregated distribution patterns typical of parasitic infections—characterized by overdispersion, wherein a minority of hosts harbor the majority of parasites—necessitate larger sample sizes to mitigate sampling bias and ensure ecological validity. As emphasized by Shvydka et al. (2018), a minimum of 80 host individuals is essential to reliably estimate parasite abundance and prevalence; smaller cohorts risk skewed interpretations, either underestimating or overestimating true infection dynamics. These findings highlight the critical need for future research to prioritize adequate sampling frameworks to ensure accurate estimations and more effective management strategies. The following ectoparasites, shown in Table 1, are recognized for their zoonotic relevance: Leptotrombidium delicense (Chigger), O. bacoti (Mites), X. cheopis (Flea), and Haemaphysalis longicornis (Ticks), (Lice). This review highlights the risk of zoonotic transmission from ectoparasites of various genera and causal agents. Leptotrombidium deliense is a medically important chigger mite and a potent vector of scrub typhus. The abundance of these species indicates a possible concern regarding the spread of this new illness in these places. Scrub typhus is caused by Orientia tsutsugamushi and is transmitted by an arthropod vector of the Trombiculidae family chigger mites L. delicense. (Paulraj et al., 2022). The mite L. delicense, which is usually known as the chigger, is the main carrier of scrub typhus. Chiggers are found in several countries in Southeast Asia, including India, Indonesia, and Thailand. These mites live in certain areas of soil called mite islands, which provide the best circumstances for them to survive. These mites’ larvae attack people in certain areas, leading to infection. (Jindal et al., 2025). The larval stage of trombiculid mites primarily transmits scrub typhus. The larvae, known as chiggers, are the only stage that actively parasitize animals or humans, making them a primary concern for transmission. Infected chiggers disseminate the disease through transovarial transmission, maintaining the bacterium within the mite population throughout many life stages. The principal reservoirs of scrub typhus pathogens are trombiculid mites, namely those of the genus Leptotrombidium (Jindal et al., 2025). According to reports, most chigger mites, including L. deliense, thrive in hot and humid settings (Chen et al., 2023). The tropical rat mite (O. bacoti), a globally prevalent species of gamasid mite, is found in almost all regions of the planet, excluding the Arctic and Antarctic areas. Ornithonyssus bacoti is the main type of gamasid mite important to medicine. All of its life stages—larvae, protonymphs, deutonymphs, and adults—can invade animals and bite them to feed on their blood (Huang et al., 2013). Dermatitis resulting from the stinging of O. bacoti mites is commonly documented globally (d’Ovidio et al., 2018). Ornithonyssus bacoti not only directly stings humans but also contributes to the transmission of certain zoonoses. O. bacoti, along with another mite species called Allodermanyssus sanguineus, is known to spread rickettsialpox, a disease caused by Rickettsia akari (Pranab et al., 2016; Sargison et al., 2025) . Rickettsialpox, a zoonotic disease linked to rodents, is mostly observed in the United States, Canada, Russia, India, Ukraine, Egypt, and the United States, among other regions (Leibler et al., 2016). In addition to being a vector for hemorrhagic fever with renal syndrome (HFRS) caused by hantavirus, it has been demonstrated to be a possible vector for HFRS induced by hantavirus (Yin et al., 2021). The potential role of O. bacoti mites as vectors for zoonotic diseases has been suggested, indicating their capacity to transmit pathogens, including endemic typhus, Bartonella spp., Q fever, tularemia, epidemic hemorrhagic fever, coxsackievirus, Yersinia pestis, and rickettsialpox (Sargison et al., 2025). Xenopsylla cheopis, also known as the oriental rat flea, serves as a significant vector for several infections. The principal vector of Yersinia pestis, the etiological agent of bubonic plague in humans, and an intermediate host for helminths, including Hymenolepis diminuta and Hymenolepis nana (Kusumarini et al., 2021). Yersinia pestis exists around the world through wildlife cycles that involve burrowing rodents and the fleas that live on them. The flea species X. cheopis is most effective at spreading the bacteria because it is often blocked in a way that helps transmit the disease (Pauling et al., 2024). Laboratory studies confirmed that infected X. cheopis fleas can transmit viable Y. pestis transovarially (from adults to eggs), enabling bacterial persistence across all subsequent flea life stages, even in the absence of detectable mammalian plague activity (Pauling et al., 2024). Additionally, X. cheopis is the sole ectoparasite capable of transmitting murine typhus via flea feces contaminated with Rickettsia typhi, which enters humans through inhalation or bite wounds (Kusumarini et al., 2021; Setiati et al., 2021). This species is also recognized as the main global vector of murine typhus, epidemic typhus, and bartonellosis, and it remains the most commonly reported flea vector in regions such as Indonesia (Manyullei et al., 2020). Haemaphysalis longicornis, commonly known as the Asian longhorned tick, is a hard tick native to eastern Asia, Australia, New Zealand, and several Pacific islands (Namjoshi et al., 2024; Okely et al., 2025). Since first reported in the United States in 2017, it has spread throughout 15 states (Okely et al., 2025). This tick species exhibits both bisexual and parthenogenetic reproduction, enhancing its invasive potential(Namjoshi et al., 2024). Haemaphysalis longicornis is a known vector of at least 30 human pathogens, including spotted fever group rickettsia, Anaplasma spp., Borrelia burgdorferi sensu lato, and Babesia spp., (Namjoshi et al., 2024). The first reported human bite occurred in 2019. In Asia, it transmits severe fever with thrombocytopenia syndrome virus (SFTSV), whereas in the United States, it has been linked to Anaplasma phagocytophilum and Heartland virus (Xu et al., 2024). Additionally, it has been found to carry emerging viruses such as Dabieshan tick virus and Langat virus (Xu et al., 2024). As a three-host tick, H. longicornis parasitizes livestock (sheep, cattle, goats, horses), wildlife (deer, rodents, birds), domestic pets, and humans (Okely et al., 2025). Its adaptability, wide host range, and reproductive flexibility facilitate rapid geographic expansion (Namjoshi et al., 2024). Migratory birds may further contribute to the spread of their presence across continents (Okely et al., 2025). Climate change, urbanization, and global trade have accelerated the worldwide spread of vector-borne diseases, including H. longicornis (Okely et al., 2025). Temperature and humidity significantly affect the reproduction and survival of Amphidinium, increasing its potential to establish in new areas (Namjoshi et al., 2024). The ability of ticks to withstand diverse climatic conditions has become a growing concern for human and animal health (Okely et al., 2025). Polyplax spinulosa, a blood-sucking louse, has been reported in 45% of rodents in Metro Manila (urban) and 100% in CALABARZON (rural). Its occurrence is influenced by environmental factors (Chan et al., 2024). Mild infestations are often undetected but can become evident if the host weakens or experiences severe stress. In rodents, symptoms include scratching, hair loss, and skin changes (Chan et al., 2024). Beyond their role as pathogen vectors, ectoparasites such as P. spinulosa cause skin irritation in mammals, potentially leading to dermatitis (Chan et al., 2024). High infestation levels weaken rats, affecting their activity and demonstrating a host–parasite relationship. Additionally, Rattus spp. serve as vectors for P. spinulosa, facilitating parasite transmission to humans, especially in impoverished communities where rats thrive (Chan et al., 2024). Lice are species-specific but can harbor zoonotic pathogens such as plague bacilli and transmit diseases such as tularemia and bartonellosis to humans. In addition, severe infestations can cause intense pain, irritation, and anemia (Farid et al., 2021). In the WHO Eastern Mediterranean Region, Bartonella rattimassiliensis has been detected in P. spinulosa and Hoplopleura pacifica in Egypt (Tahmasebi Ashtiani et al., 2024). Zoonotic endoparasites in rodents inhabiting Indonesia: prevalence, transmission, and public health implicationsWild rodents act as definitive, intermediate, and paratenic hosts for several endoparasites (helminths and protozoa) with zoonotic potential, including Capillaria hepatica, Cryptosporidium spp., Giardia spp., T. gondii, and Schistosoma spp. (Riquelme et al., 2021). Their role in disease transmission is a public health concern. Rodents also serve as hosts for various ectoparasites, such as fleas, ticks, mites, and lice, which act as disease vectors. Many intermediate hosts of diseases, including fleas and ticks, contaminate human food stores, increasing zoonotic infections in urban settings (Archer et al., 2017; Hardgrove et al., 2021; Wardhana et al., 2024b). In informal settlements and slums, where animals roam freely and open defecation is widespread, the mechanical spread of parasites by rats presents an especially high risk of disease transmission (Garedaghi and Khaki, 2014; Archer et al., 2017). Endoparasites, or intestinal parasites, inhabit the host’s gastrointestinal tract, lungs, bodily cavity, blood, or various tissues. They encompass helminths, which are multicellular organisms categorized into nematodes (roundworms), trematodes (flatworms), and cestodes (tapeworms), as well as protozoa, which are unicellular organisms that predominantly circulate in the bloodstream (Mohd-Qawiem et al., 2024). Helminths are typically found in the intestines, muscles, liver, and urinary bladder, whereas protozoa can infect a variety of tissues and organs (Mustapha et al., 2019; Mohd-Qawiem et al., 2024). Studies in Durban reported the presence of two zoonotic parasites, H. nana and Hymenolepis diminuta in Rattus norvegicus. Serological testing also revealed T. gondii antibodies in 4.1% of R. norvegicus and 35% of human inhabitants living in an informal settlement (Ogolla et al., 2019). Global research on rodent endoparasites has garnered significant attention, particularly in China (Chaisiri et al., 2015), Lao People’s Democratic Republic (Pakdeenarong et al., 2014), Malaysia (Tijjani et al., 2020), the Netherlands (Franssen et al., 2016), Iran (Garedaghi and Khaki, 2014; Shirvan et al., 2024), and Indonesia (Puspitasari et al., 2023). Eighteen published research studies on endoparasites in wild rats have been completed in Indonesia, employing diverse approaches and techniques for parasite detection (Afililla et al., 2017; Katsumata et al., 2018; Herawati and Suparmani, 2020; Resnhaleksmana et al., 2020; Mamonto et al., 2020; Resnhaleksmana et al., 2021; Kusumarini et al., 2021, 2022; Herawati and Sudarmaji, 2021; Setiati et al., 2021; Yesica et al., 2022, 2024; Puspitasari et al., 2023; Setiati and Fatmawati, 2023; Wardhana et al., 2024a; Wardhana et al., 2024b; Pratama et al., 2024; Hutagalung et al., 2024). Ten of the 18 research studies looked at the host with a rather small sample size (>80 people) (Afililla et al., 2017; Katsumata et al., 2018; Mamonto et al., 2020; Resnhaleksmana et al., 2021; Kusumarini et al., 2021, 2022; Yesica et al., 2022, 2024; Wardhana et al., 2024b; Hutagalung et al., 2024). Previous research on endoparasites showed that rats in Indonesia can be infected by five types of endoparasites: cestodes, nematodes, acanthocephalans, intestinal protozoa, and blood protozoa. The following species, among the aforementioned endoparasites, have zoonotic implications: Hymenolepis nana, Diphyllobothrium sp., Hymenolepis sp., Hymenolepis diminuta, and Taenia taeniaeformis (Cestodes); Trichuris trichiura and Strongyloides spp. (nematode); Moniliformis moniliformis (acanthocephalan); Cryptosporidium parvum, Cryptosporidium spp., T. gondii, Blastocystis (intestinal protozoa), and T. lewisi (blood protozoa). Taenia taeniaeformis (syn. Hydatigera taeniaeformis, a member of the Taeniidae family of cestodes, is a zoonotic parasite with a global distribution, particularly in areas where rodent and cat populations overlap. It primarily infects felids, such as domestic and wild cats, as definitive hosts (Islam et al., 2024), but it has also been found in canids and mustelids, while rodents (e.g., Rattus rattus, Tatera indica) serve as intermediate hosts (Herawati and Sudarmaji, 2021). The lifecycle of T. taeniaeformis begins when infected cats pass embryonated eggs in their feces, which are then ingested by rodents. The eggs hatch in the small intestine, and the larvae migrate to the liver, where they develop into cystic strobilocerci. The definitive host becomes infected upon consuming an infected rodent, thereby completing the cycle (Herawati and Sudarmaji, 2021). Although T. taeniaeformis infections in felids are generally asymptomatic, heavy infestations can lead to vomiting, anorexia, lethargy, dyspnea, and acute intestinal obstruction. In rodents, the parasite forms exudate-filled cysts in the liver and surrounding organs, contributing to morbidity. Infection rates in rodents are influenced by age, population density, seasonal variation, feline behavior, rodent community dynamics, and meteorological conditions. Given its zoonotic potential, T. taeniaeformis poses a threat to human health, especially in areas where humans have close contact with infected definitive hosts or consume contaminated food and water (Thaikoed et al., 2024). Hymenolepis diminuta has a well-defined lifecycle (egg → cysticercoid → adult), adaptability to laboratory conditions, and interactions with host physiology (e.g., microbiota modulation), making it indispensable for studying cestodiasis mechanisms and host–parasite dynamics (Sulima-Celińska et al., 2022). Despite its global distribution, human infections remain rare (0.001–5.5% prevalence), often paucisymptomatic, and linked to accidental insect consumption. Case reports from Jamaica, India, Romania, and Italy highlight the sporadic but widespread occurrence of this disease, underscoring the need for surveillance in rodent- and insect-prone areas (Galoş et al., 2022). Hymenolepis diminuta dominance in murine populations (91% single-species infections) reduces competition with other helminths, enhancing their transmission potential. This ecological advantage poses risks to cross-species spillover to humans in shared environments (Mamonto et al., 2020). Trichuris trichiura, a soil-transmitted helminth (STH), infects approximately 464 million people worldwide, contributing to 0.64 million disability-adjusted life years lost annually due to complications such as anemia, malnutrition, and Trichuris Dysentery Syndrome in children (Behniafar et al., 2024). The parasite lifecycle begins when embryonated eggs are shed in feces, mature in soil (15–30 days), and infect hosts via contaminated food or water. Adult worms colonize the large intestine, producing 3–20,000 eggs daily (Behniafar et al., 2024). Although mild infections are often asymptomatic, heavy burdens cause abdominal pain, bloody diarrhea, and growth retardation, particularly in underserved tropical regions with poor water, hygiene, and sanitation (WASH) infrastructure. Globally, 144 million preschool and 233 million school-aged children are affected, with prevalence exceeding 90% in high-risk communities (Behniafar et al., 2024). With 70.6 million cases, Indonesia has the second highest incidence of STH in the world, thirty percent of which impact preschool-age children (Djuardi et al., 2021). A 2008 national survey found that 61% of the population was infected with STHs, including over 90 million Ascaris lumbricoides and 60 million T. trichiura cases (Djuardi et al., 2021). High endemicity has been reported in Banten, Jakarta, South Sulawesi, Bali, and Papua (Djuardi et al., 2021). In East Nusa Tenggara, 19.7% of adults are infected with T. trichiura, as well as hookworms (51.7%) and A. lumbricoides (21.8%) (Djuardi et al., 2021). Local studies in North Sumatra’s Simanindo and Ronggur Nihuta districts found 4.8% and 5.9% prevalence of STH in children, respectively, with T. trichiura dominating in Simanindo (100% of cases) and co-infections occurring in Ronggur Nihuta (Ipa et al., 2024). Despite reported access to safe water, seasonal shortages during dry periods exacerbate transmission risks (Ipa et al., 2024). T. trichiura exacerbates malnutrition and anemia by feeding on host blood, particularly in children with moderate-to-severe infections (Djuardi et al., 2021; Agustina et al., 2022). Infections also alter gut microbiota composition, reducing microbial diversity and elevating Firmicutes dominance, which may impair nutrient absorption and immune function (Chen et al., 2021). Although acute infections are often asymptomatic, chronic cases can lead to fatal complications, such as colon perforation. Diagnosis relies on stool microscopy (e.g., Kato-Katz method), but asymptomatic cases complicate surveillance (Agustina et al., 2022; Behniafar et al., 2024). The high prevalence of T. trichiura and A. lumbricoides in Indonesia reflects widespread oral–fecal transmission driven by inadequate WASH infrastructure and seasonal water scarcity (Djuardi et al., 2021; Ipa et al., 2024). Targeted interventions, including deworming programs and improved sanitation, are critical for reducing transmission. However, the parasite’s resilience and the asymptomatic nature of mild infections necessitate sustained community engagement and monitoring in endemic regions. Toxoplasma gondii is a globally prevalent protozoan that causes zoonotic toxoplasmosis, affecting over a third of the population (Ahidjo et al., 2025). It spreads via oocysts excreted by cats, with wild rats serving as intermediate hosts and indicators of contamination (Puspitasari et al., 2023). The parasite progresses through sporozoite (infective), tachyzoite (immune-modulating), and bradyzoite (tissue cyst) stages. Chronic brain cysts may alter host behavior and persist for life (Wana et al., 2023). Although often asymptomatic in immunocompetent individuals, congenital and immunocompromised cases can lead to severe neurological and ocular issues (Ahidjo et al., 2025). A Surabaya study found T. gondii seroprevalence in 31% of wild rats (30% IgG, 1% IgM), with 19% carrying brain cysts. The infection risk was highest in densely populated areas (50% IgG, 2.8% IgM), which was linked to cat presence (Puspitasari et al., 2023). The global seroprevalence of T. gondii in rodents varies significantly, ranging from 0.3% to 76%, reflecting regional and species-specific differences. The reported rates are 56% in Iran (Hosseini et al., 2021), 5.5% in Poland (Grzybek et al., 2021), and 0.8% in West India (Dubey et al., 2016). Additional studies reported 76% prevalence in central rock rats in Nigeria (Ode et al.), 35% in various rodents in Iran (Nazari et al.), 11% in Rattus rattus in Saudi Arabia (Elamin), 8% in rodents in Brazil (Ruffolo et al.), 7.3% in nine rodent species in Romania (Kalmár et al.), 5.9% in five species in Malaysia (Normaznah et al.), 4% in Rattus norvegicus and R. rattus in Egypt (Mikhail et al.), and 3.6% in Mus musculus in Argentina (Salcedo et al.). These findings highlight the widespread nature of T. gondii infection among rodents and underscore the need for continued monitoring and research. Beyond rodents, T. gondii infection is also prevalent in other animals, with seroprevalence reaching up to 75% in dogs and 73% in cats, as well as in humans, where rates can be as high as 70% (Puspitasari et al., 2023). Rats acquire T. gondii through contaminated food, vertical transmission, cannibalism, and infected insects, facilitating its spread to cats and other animals. Surveillance in cats is essential for controlling transmission (Puspitasari et al., 2023). Trypanosoma lewisi is a globally distributed, obligate rodent blood parasite transmitted by fleas (X. cheopis) (Wardhana et al., 2024a; Yesica et al., 2024). Although typically non-pathogenic in rodents, this zoonotic agent causes atypical human trypanosomiasis, with sporadic and occasionally fatal human cases reported in Asia and Africa (Wardhana et al., 2024a). The parasite thrives in rodent-dense environments, with fleas facilitating rapid dissemination. Although rare, human infections highlight their zoonotic potential, necessitating surveillance in areas with rodent-flea co-existence (Wardhana et al., 2024a). Molecular techniques for parasite detection and identificationIn Indonesia, research on parasite species determination through molecular methods remains insufficient, with six published studies on rodents in the past decade (2015–2025) (Katsumata et al., 2018; Resnhaleksmana et al., 2020; Resnhaleksmana et al., 2021; Wardhana et al., 2024b; Wardhana et al., 2024a; Yesica et al., 2024). Employing molecular techniques, researchers effectively identified parasites at the species level by extracting DNA from positive samples and amplifying target genes. Most of these studies focused on T. lewisi. Molecular approaches have played a crucial role in identifying and characterizing zoonotic parasites, as demonstrated in both the initial explanation and the findings from Studies 1 to 6. Polymerase chain reaction (PCR) was the primary method for species identification, and different genetic markers were targeted for amplification. For example, Katsumata et al. (2018) amplified the SSU rDNA gene to identify Blastocystis isolates, while studies (Resnhaleksmana et al., 2020; Resnhaleksmana et al., 2021) focused on the 18S rRNA gene for Cryptosporidium and the COX1 gene for Hymenolepis nana. In studies (Wardhana et al., 2024b; Wardhana et al., 2024a; Yesica et al., 2024), the ITS1 region of T. lewisi was targeted using species-specific primers (TRYP1S/TRYP1R and LEW1S/LEW1R), demonstrating consistency in the choice of genetic markers across studies. Subsequent sequencing and phylogenetic analyses provided insights into the genetic relationships and evolutionary patterns of these parasites. For instance, Katsumata et al. (2018) identified Blastocystis ST4 in wild rodents, while Resnhaleksman et al. (2021) revealed genetic kinship between Cryptosporidium parvum isolates from rats, pigs, and humans, reinforcing its zoonotic significance. Similarly, studies (Wardhana et al., 2024a; Wardhana et al. 2024b; Yesica et al., 2024) highlighted the prevalence and genetic diversity of T. lewisi, with Study 6 confirming a 98.51% genetic similarity between Indonesian isolates and a Chinese strain. Collectively, these studies underscore the zoonotic potential of Blastocystis, Cryptosporidium, H. nana, and T. lewisi, emphasizing the importance of molecular surveillance in understanding parasite transmission dynamics. Despite these advancements, the studies did not address ectoparasites, suggesting an avenue for future research. Surveillance in wild rat populations can be enhanced using a One Health approach that integrates the veterinary, public health, and environmental sectors. This approach highlights the need for coordinated efforts to address parasitic infections, focusing on transmission dynamics between the environment, animals, and humans. Improved diagnostics using molecular tools such as PCR and sequencing enable accurate detection of T. gondii. Geospatial mapping and standardized sampling protocols help identify hotspots, while sentinel sites, community engagement, and improved water sanitation strengthen early detection and control. Environmental sampling of oocysts in soil and water, along with centralized databases for data sharing, supports coordinated responses. Expanding molecular techniques to include ectoparasitic species can further enhance our understanding of zoonotic risks and improve disease control strategies. ConclusionThis study demonstrates the importance of wild rats as zoonotic parasite reservoirs in Indonesia, emphasizing the urgent need for ongoing surveillance and molecular research. While previous research frequently used limited sample sizes, limiting their general application, our findings demonstrate the significance of molecular methods in properly detecting parasites like T. lewisi. However, the molecular characterization of other zoonotic parasites is limited. To improve disease surveillance and control, future research should use improved genetic approaches, broaden sampling across many geographic locations, and incorporate environmental analysis to better understand parasite transmission dynamics. Larger sample numbers, focused species identification, and geospatial risk mapping are all advised to inform successful intervention strategies and reduce the public health hazards associated with rodent-borne diseases. AcknowledgmentsThe authors would like to acknowledge the Airlangga Post-Doctoral Fellowship (APD) program for funding this research. Conflict of interestThe authors declare no conflict of interest. FundingThe authors are grateful to the Faculty of Veterinary Medicine, Universitas Airlangga, Indonesia, for providing the facilities that supported this study. This support was provided as part of the postdoctoral program. Author’s contributionsMA, LTS, and MY drafted the manuscript. MA and LTS revised and edited the manuscripts. MA, LTS, and MY contributed to the preparation and critical evaluation of this work. MA and MY revised the references. All writers reviewed and approved the final manuscript. Data availabilityAll references are open-access, so data can be obtained from the online web. ReferencesAfililla, Z., Suwanti, L.T., Sri, S., Sudjarwo, A., Koesdarto, S. and Plumeriastuti, H. 2017. Prevalensi Trypanosomiasis tikus (Rattus sp.) liar di banyuwangi prevalence of trypanosomiasis of wild rats (Rattus sp.) in Banyuwangi. J. Parasite Sci. 1(2):39. Agustina, K.K., Wirawan, I.M.A., Sudarmaja, I.M., Subrata, M. and Dharmawan, N.S. 2022. The first report on the prevalence of soil-transmitted helminth infections and associated risk factors among traditional pig farmers in Bali Province, Indonesia. Vet. World 15(5), 1154–1162. Ahidjo, N., Seke Etet, P.F., Ngarka, L., Maidawa Yaya, F., Ndianteng, E.W., Eyenga Nna, A.L., Meka’a Zang, L.Y., Kemmo, C., Nwasike, C.N.C., Yonkeu Tatchou, F.G., Njamnshi, W.Y., Nfor, L.N., Tsouh Fokou, P.V., Djiogue, S., Fekam Boyom, F., Ngadjui, B.T. and Njamnshi, A.K. 2025. Effects of diet and ovariectomy on Toxoplasma gondii brain infection: functional alterations and neuronal loss in rats. Brain Commun. 7(1), fcae441. Archer, C.E., Appleton, C.C., Mukaratirwa, S., Lamb, J. and Schoeman, M.C. 2017. Endo-parasites of public-health importance recovered from rodents in the Durban metropolitan area, South Africa. South. Afr. J. Infect. Dis. 32(2), 57–66. Behl, A., Nair, A., Mohagaonkar, S., Yadav, P., Gambhir, K., Tyagi, N., Sharma, R.K., Butola, B.S. and Sharma, N. 2022. Threat, challenges, and preparedness for future pandemics: a descriptive review of phylogenetic analysis based predictions. Infect. Genet. Evol. 98, 105217. Behniafar, H., Sepidarkish, M., Tadi, M.J., Valizadeh, S., Gholamrezaei, M., Hamidi, F., Pazoki, H., Alizadeh, F., Kianifard, N., Nooshabadi, M.S., Bagheri, K., Hemmati, F., Hemmati, T., Tori, N.A., Siddiq, A. and Rostami, A. 2024. The global prevalence of Trichuris trichiura infection in humans (2010–2023): a systematic review and meta-analysis. J. Infect. Public Health 17(5), 800–809. Bergamo, P.E. and Santos, D. 2014. United Nations, Department of Economic and Social Affairs, Population Division, World Urbanization Prospects: The 2014 Revision, Highlights (ST/ ESA/SER.A/352). Blasdell, K.R., Morand, S., Laurance, S.G., Doggett, S.L., Hahs, A., Trinh, K., Perera, D. and Firth, C. 2022. Rats and the city: implications of urbanization on zoonotic disease risk in Southeast Asia. Proc. Nat. Acad. Sci. 119(39), e2112341119. Bonell, A., Lubell, Y., Newton, P.N., Crump, J.A. and Paris, D.H. 2017. Estimating the burden of scrub typhus: a systematic review. PLoS Negl. Trop. Dis. 11(9):e0005838. Chaisiri, K., Siribat, P., Ribas, A. and Morand, S. 2015. Potentially zoonotic helminthiases of murid rodents from the indo-chinese peninsula: impact of habitat and the risk of human infection. Vector-Borne Zoonotic Dis. 15(1), 73–85. Chan, H., Flores, M.J., Maghirang, E.S., Arellano, B. and Chan, J.M. 2024. Zoonotic ectoparasite burden in house rats (Rattus spp.) in selected urban and rural areas of NCR and CALABARZON. J. Parasite Sci. 8(1), 37–41. Chen, H., Mozzicafreddo, M., Pierella, E., Carletti, V., Piersanti, A., Ali, S.M., Ame, S.M., Wang, C. and Miceli, C. 2021. Dissection of the gut microbiota in mothers and children with chronic Trichuris trichiura infection in Pemba Island, Tanzania. Parasites Vectors 14(1):62. Chen, Y.L., Guo, X.G., Ding, F., Lv, Y., Yin, P.W., Song, W.Y., Zhao, C.F., Zhang, Z.W., Fan, R., Peng, P.Y., Li, B., Chen, T. and Jin, D.C. 2023. Infestation of oriental house rat (Rattus tanezumi) with chigger mites varies along environmental gradients across five provincial regions of southwest China. Int. J. Environ. Res. Public Health 20(3), 62. d’Ovidio, D., Noviello, E. and Santoro, D. 2018. Prevalence and zoonotic risk of tropical rat mite (Ornithonyssus bacoti) in exotic companion mammals in southern Italy. Vet. Dermatol. 29(6), 522-e174. de Almeida, A.M.P., Sobreira, M., Leal, N.C. and Tavares, C. 2020. Does the plague still threaten us? Rev. Soc. Bras. Med. Trop. 53, e20190136. Djuardi, Y., Lazarus, G., Stefanie, D., Fahmida, U., Ariawan, I. and Supali, T. 2021. Soil-transmitted helminth infection, anemia, and malnutrition among preschool-age children in nangapanda subdistrict, Indonesia. PLoS Negl. Trop. Dis. 15(6), e0009506. Dubey, J.P., Ferreira, L.R., Alsaad, M., Verma, S.K., Alves, D.A., Holland, G.N. and McConkey, G.A. 2016. Experimental toxoplasmosis in rats induced orally with eleven strains of Toxoplasma gondii of seven genotypes: tissue tropism, tissue cyst size, neural lesions, tissue cyst rupture without reaction, and ocular lesions. PLoS One 11(5), e0156255. Elamin, M.H. 2014. Genotyping of Toxoplasma gondii from rats (Rattus rattus) in Riyadh, Saudi Arabia. Kor. J. Parasitol. 52(3), 257–261. Farid, D.S., Abouelhassan, E.M., El-Sebae, A.A., Enany, M.E. and Youssef, A.I. 2021. Ectoparasite fauna of commensal rodents collected from the North Sinai governorate—Egypt and its public health significance. Adv. Anim. Vet. Sci. 9(4), 563–570. Franssen, F., Swart, A., van Knapen, F. and van der Giessen, J. 2016. Helminth parasites in black rats (Rattus rattus) and brown rats (Rattus norvegicus) from different environments in the Netherlands. Infect. Ecol. Epidemiol. 6(1), 31413. Garedaghi, Y. and Khaki, A.A. 2014. Prevalence of gastrointestinal and blood parasites of rodents in Tabriz, Iran, with emphasis on parasitic zoonoses. Crescent J. Med. Biol. Sci. 1(1), 9–12. Grzybek, M., Antolová, D., Tołkacz, K., Alsarraf, M., Borowczyk, J.B., Nowicka, J., Paleolog, J., Biernat, B., Behnke, J.M. and Bajer, A. 2021. Seroprevalence of Toxoplasma gondii among sylvatic rodents in Poland. Animals 11(4), 1048. Hardgrove, E., Zimmerman, D.M., von Fricken, M.E. and Deem, S. 2021. A scoping review of rodent-borne pathogen presence, exposure, and transmission at zoological institutions. Prev. Vet. Med. 193, 105345. Hassell, J.M., Begon, M., Ward, M.J. and Fèvre, E.M. 2017. Urbanization and disease emergence: dynamics at the wildlife–livestock–human interface. Trends Ecol. Evol. 32(1), 55–67. Herawati, N.A. and Sudarmaji. 2020. Prevalence of infection of endoparasites and its intensity in small mammals’ liver from irrigated rice areas. Adv. Health Sci. Res. 24, 141–147. Herawati, N.A. and Sudarmaji. 2021. Diversity of rodent species and its potency as the vector for transmitting rodent borne parasitic disease in households. BIO Web Conf. 33, 10. Ho, J., Changbunjong, T., Weluwanarak, T., Hussain, S. and Sparagano, O. 2021. The pests of a pest: a systematic review of ectoparasitic fauna among synanthropic rodents in the 21st century with meta-analysis. Acta Trop. 215, 105802. Hosseini, S.A., Abediankenari, S., Amauei, A., Sarvi, S., Sharif, M., Razaei, F. and Daryani, A. 2021. Seroprevalence of Toxoplasma gondii in wild rats (Rattus rattus) in Northern Iran. Vet. Med. Int. 27, 6655696. Huang, L.Q., Guo, X.G., Speakman, J.R. and Dong, W.G. 2013. Analysis of gamasid mites (Acari: Mesostigmata) associated with the Asian house rat, Rattus tanezumi (Rodentia: Muridae) in Yunnan Province, Southwest China. Parasitol. Res. 112(5), 1967–1972. Hutagalung, S.V., Mardiani, T.H., Albar, M.Y., Kurniawan, A., Sinaga, L.A., Nasution, H., Hasibuan, P. and Wardhana, A.H. 2024. Detection of rodent-borne intestinal helminths infestation in North Sumatra, Indonesia: a potential public health concern. Public Health Indones. 10(4), 124–134. Ipa, M., Isnani, T., Girsang, V.I., Amila, Harianja, E.S., Purba, Y., Wandra, T., Budke, C.M. and Purba, I.E. 2024. Soil-transmitted helminth infections and anemia in children attending government run schools on Samosir Island, Indonesia. Parasite Epidemiol. Control 25, e00344. Islam, M.M., Farag, E., Hassan, M.M., Enan, K.A., Mohammadi, A., Aldiqs, A.K., Alhussain, H., Al Musalmani, E., Al-Zeyara, A.A., Al-Romaihi, H., Yassine, H.M., Sultan, A.A., Bansal, D. and Mkhize-Kwitshana, Z. 2024. Rodent-borne parasites in Qatar: a possible risk at the human-animal-ecosystem interface. One Health 18, 100708. Issae, A., Chengula, A., Kicheleri, R., Kasanga, C. and Katakweba, A. 2023. Knowledge, attitude, and preventive practices toward rodent-borne diseases in Ngorongoro district, Tanzania. J. Public Health Afr. 14(6), 2385. Jena, S., Chawla, S. and Parthasarathy, S. 2017. Identification and morphological characterisation of spiny rat louse (Polyplax spinulosa) from a laboratory rat. J. Entomol. Zool. Stud. 5(6), 82–84. Jindal, N., Abhishek, K., Kumar, A., Topno, R.K., Pandey, K. and Sahoo, G.C. 2025. A Comparative analysis of Orientia tsutsugamushi (Scrub Typhus): essential information on causes, symptoms, and care. Clin. Microbiol. Newsl. 50, 34–40. Kalmár, Z., D’Amico, G., Mărcuţan, D.I., Paştiu, A.I., Mihalca, A.D. and Sándor, A.D. 2023. Toxoplasma gondii in small mammals in Romania: the influence of host, season and sampling location. BMC Vet. Res. 19(1), 177. Kang, C., Liu, Y. and Tong, Z. 2022. Effects of drawdown on growth and reproduction of submerged macrophyte Vallisneria spinulosa. J. Freshw. Ecol. 37(1), 285–295. Katsumata, M., Yoshikawa, H., Tokoro, M., Mizuno, T., Nagamoto, T., Hendarto, J., Asih, P.B.S., Rozi, I.E., Kimata, I., Takami, K. and Syafruddin, D. 2018. Molecular phylogeny of Blastocystis isolates from wild rodents captured in Indonesia and Japan. Parasitol. Res. 117(9), 2841–2846. Kusuma, W.A., Yudhana, A., Andriyani, A., Oktaviana, V. and Chaerunisa, N.A. 2020. First Report of Polyplax spinulosa in Albino Rats in Banyuwangi, East Java, Indonesia. J. Medik Vet. 3(1), 123–126. Kusumarini, S., Danuarta, M.F., Karami, F., Yesica, R., Wisesa, I.B.G.R., Yudhana, A. and Nurhidayah, N. 2022. Screening Strongyloides spp. infection from wild rodents implications for public awareness and attitudes on zoonotic diseases in Malang City, Indonesia. J. Medik Vet. 5(2), 196–206. Kusumarini, S., Yesica, R., Wisesa, I.B.G.R., Hermanto, J., Nurholizah, Y. and Trinastuti, M.W. 2021. Preliminary study: detection of Ecto and endoparasites among wild rats from urban area in Blimbing, Malang, East Java. Acta Vet. Indones. 2021, 95–101. Lau, A.C.C., Qiu, Y., Moustafa, M.A.M., Nakao, R., Shimozuru, M., Onuma, M., Mohd-Azlan, J. and Tsubota, T. 2020. Detection of borrelia burgdorferi sensu lato and relapsing fever borrelia in feeding ixodes ticks and rodents in Sarawak, Malaysia: new geographical records of borrelia yangtzensis and borrelia miyamotoi. Pathogens 9(10), 1–18. Leibler, J.H., Zakhour, C.M., Gadhoke, P. and Gaeta, J.M. 2016. Zoonotic and vector-borne infections among urban homeless and marginalized people in the United States and Europe, 1990-2014. Vector-Borne Zoonotic Dis. 16(7), 435–444. Mairawita, Mursyid, A., Dahelmi, Diniyati, F., Lidia, D., Putri, N., Arifa, M.M., Jefrial and Maulana, R.M. 2023. Co-occurrence of ectoparasites on wild rodents in Sipora Island, Mentawai, Indonesia with the zoonotic potential review. Biodiversitas 24(11), 6369–6376. Mamonto, H., Manyullei, S., Hamid, F., Daud, A., Syam, A. and Birawida, A. 2020. Relationship between waste with ectoparasites and endoparasites (nematodes and cestodes) in Rats. South Asian Res. J. Humanit. Soc. Sci. 2(4), 79–85. Manabella Salcedo, I., Díaz, J., Paoletti, B., Mirabella, P. and Oscherov, E. 2021. Role of Mus musculus in the transmission of several pathogens in poultry farms. Int. J. Parasitol. Parasit. Wildl. 14, 130–136. Manyullei, S., Natsir, M.F. and Batkunda, A. 2020. Identification of rat density and ectoparasites in seaport area of Manokwari, Papua Province. J. Med. Sci. 8, 204–208. Materu, A.E., Ndunguru, M.C. and Ngayabosha, N.J. 2022. A survey on ectoparasites and endoparasites of African pygmy hedgehogs (Atelerix albiventris) in urban and suburban areas of Morogoro, Tanzania. J. Entomol. Zool. Stud. 10(5), 221–225. Mikhail, M.W., Hasan, A.H., Allam, A. and Mohammed, N.M.K. 2017. Seroprevalence of Toxoplasma gondii among commensal rodents from Giza Governorate, Egypt. J. Egypt. Soc. Parasitol. 47(1), 145–150. Mohd-Qawiem, F., Nur-Mahiza, M.I., Ahmad, N.I., Yasmin, A.R. and Nur-Fazila, S.H. 2024. Zoonotic importance of ecto-and endoparasites in rodents in Malaysia: a review. J. Vet. Sci. 25(6), e81. Mustapha, T., Unyah, N.Z., Majid, R.A., Abdullahi, S.A. and Wana, N.M. 2019. Prevalence of ectoparasitic infection of rodents captured near student hostels: zoonotic implications. Annu. Res. Rev. Biol. 32(1), 1–10. Namjoshi, P., Lubembe, D.M., Sultana, H. and Neelakanta, G. 2024. Antibody-blocking of a tick transporter impairs Anaplasma phagocytophilum colonization in Haemaphysalis longicornis ticks. Sci. Rep. 14(1), 9003. Nazari, N., Armand, B., Sarvi, S., Daryani, A., Hosseininejad, Z., Moosazadeh, M. and Sharif, M. 2020. Serological survey of Neospora caninum and Toxoplasma gondii co-infection in rodents in Northwestern Iran. Iran. J. Parasitol. 15(2), 253–258. Nieto, N.C., Khan, K., Uhllah, G. and Teglas, M.B. 2012. The emergence and maintenance of vector-borne diseases in Khyber Pakhtunkhwa and tribal areas of Pakistan. Front. Physiol. 3, 250. Normaznah, Y., Noor-Ismalina, H., Azian, A.L., Zurainee, M.N. and Norlida, M.R. 2015. Seroprevalence of Toxoplasma gondii in rodents from various locations in Peninsular Malaysia. Southeast. Asian J. Trop. Med. Public Health 46(3), 388–395. Ode, S., Tersoo, D. and Musa, M.T. 2022. High prevalence of Toxoplasma gondii in Nigerian wild rats by molecular detection. Vet. Parasitol. Reg. Stud. Rep. 35, 100776. Ogolla, F.O., Omondi, C. and Odhiambo, C. 2019. Assessment of wild rodent endoparasites in Kirimiri Forest, Kenya. Int. J. Adv. Res. Publ. 3(5), 9. Okely, M., Chen, Z., Adly, E. and Kamal, M. 2025. Climate change influences on the potential geographic distribution of Haemaphysalis longicornis. Sci. Rep. 15(1), 2266. Pakdeenarong, N., Siribat, P., Chaisiri, K., Douangboupha, B., Ribas, A., Chaval, Y., Herbreteau, V. and Morand, S. 2014. Helminth communities in murid rodents from Laos: role of habitat and season. J. Helminthol. 88(3), 302–309. Palomba, M., Marchiori, E., Tedesco, P., Fioravanti, M., Marcer, F., Gustinelli, A., Aco-Alburqueque, R., Belli, B., Canestrelli, D., Santoro, M., Cipriani, P. and Mattiucci, S. 2023. An update on helminth endoparasites in the Mediterranean Sea. Parasitol. 150(12), 1139–1157. Pauling, C.D., Beerntsen, B.T., Song, Q. and Anderson, D.M. 2024. Transovarial transmission of Yersinia pestis in Xenopsylla cheopis. Nat. Commun. 15(1), 7266. Paulraj, P.S., Govindarajan, R., Ranganathan, K., Veeramanoharan, R. and Kumar, A. 2022. Ectoparasites diversity on rodents and shrews in scrub typhus-endemic areas. J. Arthropod-Borne Dis. 16(1), 51–60. Pratama, P.G., Suprihati, E., Suwanti, L.T., Hastutiek, P., Setiawan, B., Puspitasari, H. and Indasari, E.N. 2024. Detection of Cryptosporidium spp. in wild rats in Surabaya, Indonesia. Med. Kedokt. Hewan. 35(2), 71–78. Puspitasari, H., Suwanti, L.T., Mufasirin, M., Kusnoto, K., Yudaniayanti, I.S. and Setiawan, B. 2023. Tissue cysts and toxoplasmosis detection in wild rats from Surabaya, Indonesia. Open Vet. J. 13(11), 1443–1450. Resnhaleksmana, E., Artama, W.T., Wijayanti, M.A. and Fihiruddin, F. 2020. Molecular analysis of Cryptosporidium and Hymenolepis in rats from Lombok Island, Indonesia. Int. J. One Health. 6(2), 123–127. Resnhaleksmana, E., Wijayanti, M.A. and Artama, W.T. 2021. A potential zoonotic parasite: cryptosporidium parvum transmission in rats, pigs and humans in west lombok, indonesia. Afr. J. Infect. Dis. 15(2), 44–51. Riquelme, M., Salgado, R., Simonetti, J.A., Landaeta-Aqueveque, C., Fredes, F. and Rubio, A.V.2021. Intestinal helminths in wild rodents from native and exotic forests in Chile. Animals. 11(2), 1–10. Ruffolo, B.B., Silva, R.C., Silva, R.J., Garcia, J.L., Gennari, S.M. and Mineo, T.W. 2016. Isolation and genotyping of Toxoplasma gondii in seronegative urban rats and presence of antibodies in communicating dogs in Brazil. Rev. Inst. Med. Trop. Sao Paulo 58, 28. Salas, L.M., Espinoza-Carniglia, M., Schmeisser, N.L. et al. 2019. Fleas of black rats (Rattus rattus) as reservoirs of Bartonella spp. in Chile. PeerJ 2019(8), e7371. Sargison, N.D., Chaudhry, U., Costa-Junior, L., Kutcher, J.R., Li, K., Sargison, F.A. and Zahid, O. 2025. The diagnosis and vector potential of Ornithonyssus bacoti tropical rat mites in northern Europe. Vet. Parasitol. Reg. Stud. Rep. 58, 101204. Setiati, N. and Fatmawati, L. 2023. Distribution of rats and endoparasites zoonoses risk in Tandang, Semarang City. Int. J. Sci. Res. Updat. 5(1), 152–159. Setiati, N., Auliya, R., Partaya, Bodijantoro, F.P.M.H., Indriyanti, D.R. and Widiyaningrum, P. 2021. Types of Rats and Their Parasites That Potential to Transmit Disease in Tugu District, Semarang City. Biosaintifika 13(3), 363–368. Shirvan, S.P., Yaghfoori, S., Mahmoudi, A, Naddaf SR, Molawi G, Ahmadi A. 2024. Prevalence of helminths in wild rodents of Northwestern Iran. Arch. Razi Inst. 79(1), 120–128. Shvydka, S., Sarabeev, V., Estruch, V.D. and Cadarso-Suárez, C. 2018. Optimum sample size to estimate mean parasite abundance in fish parasite surveys. Helminthologia 55(1), 52–59. Sianturi, L.A., Hutagalung, S.V., Darmadi and Lubis, S.M. 2024. Identification of Mites in Wild Rats in Simpang Limun Traditional Wet Market Area, Medan -Indonesia. J. Endocrinol. Trop. Med. Infect. Dis. (JETROMI) 6(2), 54–61. Tahmasebi Ashtiani, Z., Ahmadinezhad, M., Bagheri Amiri, F. and Esmaeili, S. 2024. Geographical distribution of Bartonella spp in the countries of the WHO Eastern Mediterranean Region (WHO-EMRO). J. Infect. Public Health 17(4), 612–618. Thaikoed, S., Mitsuwan, W., Chaisiri, K., Tanthanathipchai, N., de Lourdes Pereira, M., Paul, A.K., Nissapatorn, V. and Saengsawang, P. 2024. The infection of Cysticercus fasciolaris in natural rats (Rattus species) residing in human residence areas, Nakhon Si Thammarat, Thailand. Comp. Immunol. Microbiol. Infect. Dis. 107, 102152. Tijjani, M., Majid, R.A., Abdullahi, S.A. and Unyah, N.Z. 2020. Detection of rodent-borne parasitic pathogens of wild rats in Serdang, Selangor, Malaysia: a potential threat to human health. Int. J. Parasitol. Parasites Wildl. 11, 174–182. Wana, M.N., Watanabe, M., Chiroma, S.M., Unyah, N.Z., Abdullahi, S.A., Nordin, S., Basir, R., Mohd Moklas, M.A. and Majid, R.A. 2023. Toxoplasma gondii induced cognitive impairment in rats via dysregulation of dopamine receptors and indoleamine 2,3 dioxygenase. Heliyon 9(3), e14370. Wardhana, A., Putra, F., Yudhana, A., Sawitri, D., Wiedosari, E., Mujiyanto, M., Priyambodo, S., Mufasirin, M., Hamid, P., Nugraheni, Y., Awaludin, A., Priyono, P., Dargantes, A. and Matsubayashi, M. 2024a. Detection of Trypanosoma lewisi from rodents residing in the densely populated residential regions along the coastal areas of Banyuwangi Sub District, Indonesia. Open Vet. J. 14(8), 1808. Wardhana, A.H., Sawitri, D.H., Wiedosari, E., Susanti, Mulyadi, A., Kurniawan, A., Sinaga, L.A., Hasibuan, P., Nasution, H., Rachmawati, F., Hamid, P.H. and Ristiyanto. 2024b. Molecular detection of Trypanosoma lewisi in rodents distributed in dairy cattle pens and residential areas. IOP Conf. Ser. Earth Environ. Sci. 1292(1), 012038. Widayati, A.N., Nurjana, M.A., Ardanto, A., Ristiyanto, R., Dhewantara, P.W. and Wardhana, A.H. 2020. the potential of rats and bats as reservoirs of leptospirosis and Japanese encephalitis (JE) in Muna Region, Southeast Sulawesi Province, Indonesia. Glob. J. Health Sci. 12(13), 125. Xu, X., Gao, Z., Wu, Y., Yin, H., Ren, Q., Zhang, J., Liu, Y., Yang, S., Bayasgalan, C., Tserendorj, A., Yang, X. and Chen, Z. 2024. Discovery and vertical transmission analysis of Dabieshan Tick Virus in Haemaphysalis longicornis ticks from Chengde, China. Front. Microbiol. 15, 1365356. Yeh, H.Y., Chen, K.H. and Chen, K.T. 2018. Environmental determinants of infectious disease transmission: a focus on one health concept. Int. J. Environ. Res. Public Health 15(6), 1183. Yesica, R., Hardian, A.B., Oktanella, Y., Kusumarini, S. and Prasetyo, G.J.G. 2024. Phylogenetic and Genetic Variation Analysis of ITS1 Gene of Trypanosoma lewisi in wild rats using polymerase chain Reaction. J. Medik Vet. 7(2), 335–348. Yesica, R., Shelly, K.R., Hardian, A.B., Paramanandi, D.A., Asmara, N.W., Anggara, J.E., Atmaja, P. and Gabriela Jeanne, G.P. 2022. Morphological and morphometric analysis of Trypanosoma lewisi and Toxoplasma gondii in Malang City, Indonesia Rats. J. Exp. Biol. Agric. Sci. 10(5), 1121–1128. Yin, P.W., Guo, X.G., Jin, D.C., Fan, R., Zhao, C.F., Zhang, Z.W., Huang, X.B. and Mao, K.Y. 2021. Distribution and host selection of tropical rat mite, ornithonyssus bacoti, in Yunnan Province of Southwest China. Animals 11(1), 1–15. Younis, A.E.E., Rizk, A.M. and Yassin, E.M.A. 2017. Occurence of the ectoparasites infesting Rattus rattus and R. norvigicus at El-Menofia Governorate, Egypt. Egypt. Acad. J. Biol. Sci. A, Entomol. 10(2), 65–71. Zhang, K., Fu, Y., Li, J. and Zhang, L. 2022. Public health and ecological significance of rodents in Cryptosporidium infections. One Health 14, 100364. https://doi.org/10.1016/j.onehlt.2021.100364 | ||
| How to Cite this Article |
| Pubmed Style Abuzahra M, Suwanti LT, Yunus M. A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Vet. J.. 2025; 15(6): 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 Web Style Abuzahra M, Suwanti LT, Yunus M. A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. https://www.openveterinaryjournal.com/?mno=241863 [Access: January 25, 2026]. doi:10.5455/OVJ.2025.v15.i6.6 AMA (American Medical Association) Style Abuzahra M, Suwanti LT, Yunus M. A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Vet. J.. 2025; 15(6): 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 Vancouver/ICMJE Style Abuzahra M, Suwanti LT, Yunus M. A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Vet. J.. (2025), [cited January 25, 2026]; 15(6): 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 Harvard Style Abuzahra, M., Suwanti, . L. T. & Yunus, . M. (2025) A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Vet. J., 15 (6), 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 Turabian Style Abuzahra, Mutasem, Lucia Tri Suwanti, and Muchammad Yunus. 2025. A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Veterinary Journal, 15 (6), 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 Chicago Style Abuzahra, Mutasem, Lucia Tri Suwanti, and Muchammad Yunus. "A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review." Open Veterinary Journal 15 (2025), 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 MLA (The Modern Language Association) Style Abuzahra, Mutasem, Lucia Tri Suwanti, and Muchammad Yunus. "A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review." Open Veterinary Journal 15.6 (2025), 2329-2342. Print. doi:10.5455/OVJ.2025.v15.i6.6 APA (American Psychological Association) Style Abuzahra, M., Suwanti, . L. T. & Yunus, . M. (2025) A decade of research on ectoparasites and endoparasites in wild rats in Indonesia (2015–2025): A review. Open Veterinary Journal, 15 (6), 2329-2342. doi:10.5455/OVJ.2025.v15.i6.6 |

